El Cuerpo Lúteo, nuevos mecanismos de regulación y su asociación con la infertilidad
- Inicio
- Comité Editorial
- Lineamientos
- Carta de Cesión de Derechos
- Información Legal
- Acerca de la Revista
- Bases de Datos
- Contacto
- ISSN 2007-3054
- Centro de Investigaciones Cerebrales
Universidad Veracruzana
Bravo-Benítez Juan Manuel1, Medel Rojas Alfonso2, Mirto-Aguilar Nancy2, Cruz Gómez Yolanda3, Morán Raya Carolina2*
1Doctorado en Ciencias Biológicas, Universidad Autónoma de Tlaxcala, Tlaxcala, México. 2Instituto de Ciencias, Benemérita Universidad Autónoma de Puebla, Puebla, México. 3Centro Tlaxcala de Biología de la Conducta, Universidad Autónoma de Tlaxcala, Tlaxcala, México.
Resumen
Abstract
Introducción
Duración del Cuerpo Lúteo
Fenómenos de transición y luteinización
Angiogénesis
Mecanismos de regulación del Cuerpo Lúteo
Regresión Lútea
Inervación y Cuerpo Lúteo
Inervación y Cuerpo Lúteo
Catecolaminas y CL
La insuficiencia de la Fase Lútea
Conclusiones
Conflicto de intereses
Agradecimientos
Referencias
Mail
El cuerpo lúteo es un tejido dinámico que se forma después de que el ovocito es liberado del folículo ovárico. Su principal función es la producción de progesterona para el establecimiento y mantenimiento de la preñez. En esta revisión se abordan temas relacionados con la formación, función y regresión del cuerpo lúteo. Se describen procesos fisiológicos tales como angiogénesis y el efecto de hormonas no esteroideas producidas por el cuerpo lúteo. También se incluye una sección sobre inervación intrínseca del ovario, incluyendo al cuerpo lúteo, un tema controvertido ya que se considera que esta estructura no recibe inervación. Los avances en la comprensión de la fisiología del cuerpo lúteo ayudarán a mejorar nuestro conocimiento sobre la fisiología de las gónadas, fundamentalmente para buscar nuevos tratamientos a problemas de fertilidad asociados a insuficiencia lútea, por lo que es altamente relevante tanto para la clínica humana como en la zootecnia.
Palabras clave: Inervación ovárica, ovulación, Función ovárica, Cuerpo Lúteo, Insuficiencia Lutea.
The corpus luteum is a dynamic tissue that is generated after an oocyte is released by an ovarian follicle. It’s main function is the production of progesterone for the establishment and maintenance of pregnancy. This review addresses issues related to the formation, function, and regression of the corpus luteum. Physiological processes such as angiogenesis and the effect of non-steroidal hormones produced by the corpus luteum are described. The intrinsic innervation of the ovaries, including the corpus luteum is also introduced, a controversial topic since this structure is considered as non-innervated. Advances in the understanding of the physiology of the corpus luteum will help to improve our knowledge about the physiology of the gonads, primarily focusing on new treatments for fertility disorders related to luteal insufficiency. This information is highly relevant for both human clinical and livestock farming areas.
Keywords: Ovarian Innervation, Ovulation, Ovarian Function, Corpus Luteum, Luteal Insufficiency.
La supervivencia de las especies depende del proceso reproductivo, en el caso de los mamíferos el funcionamiento cíclico de los ovarios es indispensable, el cual de manera general consiste en una fase folicular y una fase lútea. El ovario es regulado por el eje hipotálamo-hipófisis-gónadas mediante sistemas de retroalimentación positiva y negativa. El Cuerpo Lúteo (CL) es un tejido dinámico que se forma del tejido remanente del folículo de Graaf, una vez que el ovocito ha sido liberado del folículo ovárico. Su función principal es la producción de progesterona para el establecimiento y mantenimiento de la preñez. En esta revisión se describe que el papel del CL no solamente es endócrino, más bien se muestra como una estructura compleja y dinámica que interacciona con otros sistemas. También se ponen en evidencia las áreas que deben estudiarse para conocer sobre sus funciones y posibles campos por abordar sobre problemas de índole clínico relacionados con la disfunción del CL.
TDe acuerdo a la vida media del CL posterior a la ovulación y donde no hubo fecundación, los mamíferos pueden ser divididos en tres categorías: (A) de CL de larga duración, (B) de CL de corta duración y (C) de CL de ultra-corta duración. A las especies que tienen CL de larga duración se les conoce como monoéstricas estacionales, porque tienen un ciclo estral anual. Incluye a carnívoros (gatos, perros, lobos, zorros, hurones y zorrillos) y otros animales como el ciervo, armadillo y marsupiales, cuya duración del CL varía entre dos semanas y seis meses.1 En la hembra de Canis Familiaris, la duración de la fase lútea es de alrededor de 65 días se encuentre preñada o no preñada.
Los animales con CL de corta duración son individuos con varios ciclos estrales al año (especies poliéstricas), cuyo CL se desarrolla y funciona en un intervalo finito de tiempo durante el ciclo ovárico, sin embargo, la vida del CL incrementa marcadamente en la preñez. Las especies de esta categoría incluyen a los primates (monos, grandes simios y humanos), en los que después de la ovulación se forma un CL que es funcional durante dos semanas para permitir el movimiento del embrión a través del cuello uterino y para preparar al útero para la implantación. Por último, los animales con CL de ultra-corta duración, son especies poliéstricas que no forman un CL funcional (roedores poliéstricos y algunos insectívoros), incluye también a los ovuladores reflejos, como la coneja, aquellos donde el apareamiento provoca la ovulación o pseudopreñez.2
3. Fenómenos de transición y luteinización
La regulación de la esteroidogénesis del CL puede desglosarse en tres acontecimientos importantes; luteinización (conversión de un folículo ovulatorio), regresión lútea y mantenimiento/rescate de la preñez. Los factores que controlan estos eventos y dictaminan su variabilidad en las diferentes especies de mamíferos dependen de la composición de las células lúteas grandes (LLC, por sus siglas en inglés) derivadas de las células de la granulosa del folículo, de las células lúteas pequeñas (SLC, por sus siglas en inglés) derivadas de las células foliculares de la teca y de las enzimas implicadas en la vía esteroidogénica, aunque estas últimas son relativamente similares entre las especies de mamíferos.3
La transición de un folículo preovulatorio a un CL es un proceso con mecanismos complejos similares a la cicatrización de heridas y formación de tumores, básicamente en la obtención de un suministro adecuado de nutrientes al acceder al sistema vascular del órgano.4,5 Antes de la ovulación, se presentan cambios estructurales en las células de la granulosa y de la teca. Por ejemplo, en el núcleo, la cromatina se dispersa y se forma el nucléolo, lo que va acompañado de un aumento en el número de polirribosomas en el citoplasma. Las uniones comunicantes entre las células de la granulosa y también entre las células de la teca desaparecen, presumiblemente como un preludio a la remodelación celular. A medida que las células se redondean y sus crestas cambian de forma laminar a tubular, la cantidad de retículo endoplásmico liso de las células incrementa y se modifican las características estructurales mitocondriales.5 Aunado a la remodelación tisular se interrumpe la proliferación celular presentándose hipertrofia y diferenciación de células esteroidogénicas del folículo, lo que da lugar a las células lúteas del CL. Al mismo tiempo, en las capas celulares de la granulosa del folículo que previamente eran avasculares se presenta un proceso de formación acelerada de nuevos vasos sanguíneos (angiogénesis). En algunas especies también se forman nuevos vasos linfáticos (linfatogénesis). En el CL la angiogénesis permite la formación de una base microvascular extensa. 6
Aunque se cree que la formación rápida del CL postovulación se debe principalmente a un proceso de hipertrofia (aumento en el tamaño del volumen de células) más que a un proceso de hiperplasia (aumento en el número de células), durante el desarrollo del CL de bovinos se ha observado proliferación de células SLC, lo que sugiere que el CL se forma por los dos procesos; hipertrofia e hiperplasia (Figura 1). La capacidad proliferativa de las SLCs disminuye conforme se pasa del estadio de crecimiento a la etapa de desarrollo y regresión del CL. 7
La angiogénesis fisiológica en adultos es un evento raro, con pocas excepciones como la vasculogénesis necesaria para el crecimiento de tejido en la cicatrización o en los órganos reproductores femeninos, como el útero y el ovario. La angiogénesis en el CL tiene su origen en la vasculatura del folículo en desarrollo. Previo a la ovulación, la membrana basal del folículo impide la entrada de los vasos sanguíneos a la capa de células de la granulosa. Cuando el folículo colapsa, lo efectúa hacia adentro, y la capa de células de la teca junto con sus vasos sanguíneos se sitúan dentro de los pliegues del compartimento granuloso. Con la pérdida de la integridad de la membrana basal se presentan extensas remodelaciones tisulares, dando comienzo a la invasión vascular en la región que contiene las células luteínicas y el subsecuente desarrollo de nuevos vasos sanguíneos a partir de la vasculatura preexistente. La sangre y el plasma se extravasan en la cavidad folicular donde forman un coágulo rico en fibrina, originando una serie de eventos subsecuentes que se asocian con un período de intensa angiogénesis donde la vasculatura formada se extiende a través del tejido 6,7
Particularmente en el CL, la regulación del proceso angiogénico parece estar controlada por acciones resultantes del equilibrio entre factores pro- y antiangiogénicos.8 Aunque poco se conoce sobre los mecanismos celulares específicos involucrados, en general se plantea que el factor de crecimiento endotelial vascular (VEGF-α) producido por las células lúteas tempranas, actúa de forma paracrina a través de receptores/correceptores de VEGF (por ejemplo, neuropilinas) en células endoteliales microvasculares para promover la angiogénesis. Además, los factores locales, como los factores de crecimiento similares a la insulina (IGF) -1 y -2, pueden generar sinergias con la hormona luteinizante (LH) para promover la producción de VEGF,9 en el CL de primates el VEGF actúa inicialmente como un factor angiogénico, pero posteriormente como un factor trófico (Figura 2).10-12
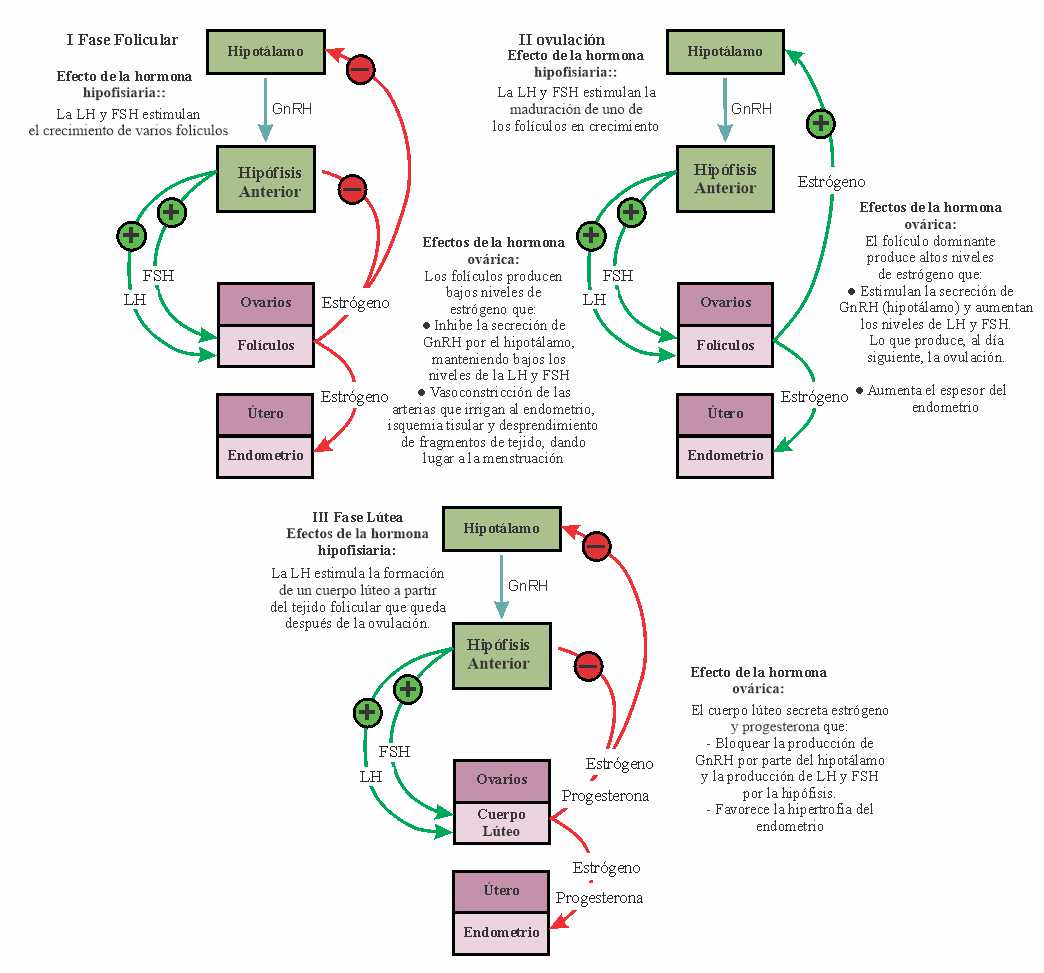
Figura 1. ARegulación hormonal de los ciclos ováricos y menstruales mediante sistemas de retroalimentación positiva y negativa en el eje hipotálamo-hipófisis-gónadas.8
Por otro lado, se ha observado que los esteroides ováricos influyen en el volumen de irrigación del tejido y la sensibilidad a los receptores β-adrenérgicos: el bloqueo de estos receptores provoca un aumento en el flujo sanguíneo ovárico, mientras que el bloqueo de los α-receptores causa el efecto opuesto. Es decir, que la noradrenalina (NA), a través de la unión con sus receptores (α o β-adrenérgicos), aumenta el flujo de sangre de los ovarios hasta en un 60%. El flujo de sangre ovárica en ovinos y bovinos es más alto durante la fase lútea media y notablemente más bajo durante el estro. La presencia de un CL funcional parece estar asociada con una capacidad vasodilatadora local pronunciada, ya que aproximadamente el 90% del flujo de sangre a los ovarios es influenciada por el CL durante la fase lútea media. Además de esto, la progesterona reduce la tasa de degradación de NA al inhibir la actividad de la mono-aminooxidasa (MAO) y la catecol-o-metiltransferasa (COMT), por lo tanto, al aumentar la vida media del neurotransmisor puede suponerse que el ovario, por medio de sus propios esteroides, regula el flujo de sangre a través del tracto reproductivo.13
Teniendo en cuenta las etapas tempranas de la formación del CL y su importancia en la preñez temprana, este mecanismo protege y apoya la función normal del CL. Sin embargo, aún no están bien esclarecidos los mecanismos precisos mediante los cuales aparece la neovascularización lútea, puede ser a través de la angiogénesis (expansión de los vasos sanguíneos existentes) o posiblemente mediante la vasculogénesis (por ejemplo, de las células progenitoras vasculares circulantes) que se forman la densa red capilar en el tejido lúteal diferenciado.1,7,8
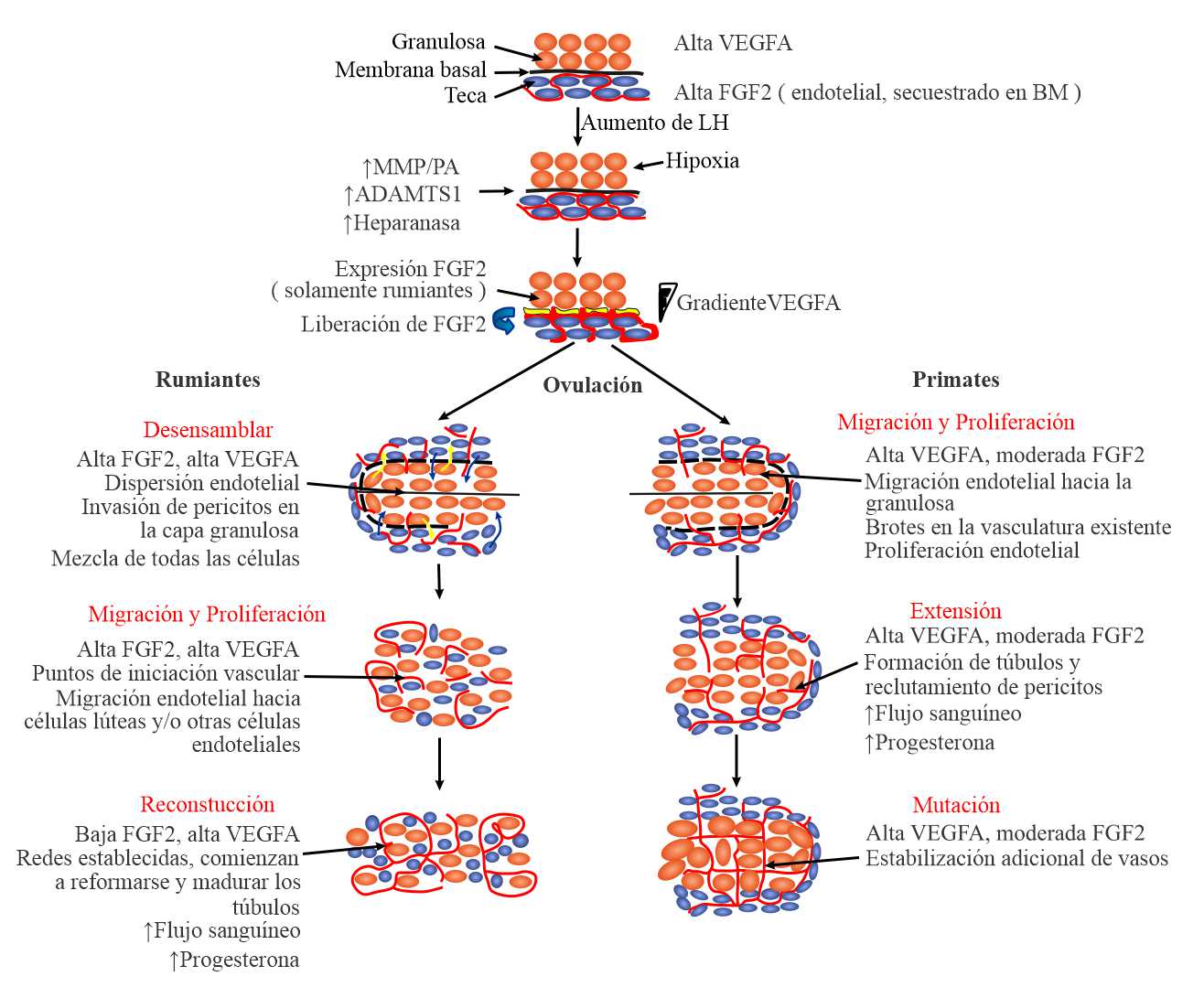
Figura 2. Mecanismos de vascularización del CL en rumiantes y primates. Las células de la granulosa y la teca se muestran en naranja y azul respectivamente, la vasculatura en rojo (no se distinguen). En el folículo preovulatorio, la capa granulosa permanece avascular, mientras que la teca presenta una extensa vascularización.7
5. Mecanismos de regulación del Cuerpo Lúteo
La formación y duración limitada del CL son críticas para la fertilidad, ya que la progesterona es la hormona esteroide esencial requerida para la implantación del embrión y mantenimiento del embarazo intrauterino hasta que se desarrolla la placenta. La actividad lútea depende de la interacción de diversos factores de crecimiento, citocinas y hormonas, incluyendo las hormonas tiroideas.14 De los varios factores luteotróficos, tres hormonas juegan un papel principal que, dependiendo de la especie, actúa para funcionar individualmente o como parte de un complejo: LH, Prolactina (PRL) y Estradiol (E2).15 El aumento preovulatorio de las gonadotropinas (LH, Hormona Folículo Estimulante [FSH, por sus siglas en inglés]) y de PRL promueven la diferenciación morfológica de las células foliculares y está asociada con los cambios bioquímicos involucrados en la luteinización. En roedores, la PRL es luteotrófica durante el periodo de preñez, pero luteolítica durante los ciclos no fértiles. La PRL estimula la angiogénesis actuando directamente en las células endoteliales o indirectamente mediante la estimulación de factores proangiogénicos como el VEGF. Por otra parte, las propiedades antiangiogénicas de PRL se presentan después de una escisión proteolítica por vasoinhibinas, una familia de fragmentos de PRL (incluida la PRL de 16 kDa) con potentes efectos antiangiogénicos, vasoconstrictores y en contra de la permeabilidad de los vasos.16
En la mujer, la LH y la hormona coriónica humana (hCG), son las hormonas luteotrópicas vitales durante el ciclo menstrual y el embarazo temprano, respectivamente. En lo que se refiere a los mecanismos celulares y endocrinos que regulan el funcionamiento del CL en humanos y en otras especies de mamíferos, se tienen puntos en común y algunas diferencias importantes. Por ejemplo, los primates y bovinos tienen un contenido similar de células LLC y SLC, y un gran número de células endoteliales capilares que forman la vasculatura, las cuales tienen un papel esencial en la función óptima del CL. Cabe destacar que las células lúteas en rumiantes crecen más que en los primates y adquieren una capacidad para producir progesterona que es independiente de la estimulación de LH. A diferencia del CL de los bovinos, las LLC de los primates siguen requiriendo la estimulación por LH y la hCG a lo largo de la fase lútea. Aunque los folículos preovulatorios de mujeres y vacas son de tamaño similar, así como la producción esteroidogénica (10 a 20 mg/h), el CL bovino produce aproximadamente diez veces más progesterona en comparación con el CL humano (17.4 mg/h frente a 1.4 mg/h), posiblemente debido al mayor desarrollo de las células lúteas bovinas que producen la hormona.17
En los primates y los rumiantes, el CL es en gran medida, dependiente de la LH que actúa a través del AMPc/proteína cinasa. En contraste, en roedores y conejos, la PRL y el E2 son críticos como factores de la luteólisis.3 En la mujer, el patrón de secreción de LH y la producción de progesterona tienen las siguientes características: (a) la frecuencia de liberación pulsátil de la LH disminuye progresivamente correlacionándose con la duración de la exposición a progesterona; (b) la amplitud de los pulsos de LH varía con la aparición de una mayor cantidad de pulsos más pequeños que se correlacionan bien con el nivel de progesterona; (c) en la fase lútea temprana, el patrón de secreción de progesterona es estable; (d) en el centro y fase lútea tardía, la secreción de progesterona es episódica, y se correlaciona con la liberación pulsátil de LH; y (e) la progesterona sola en la fase lútea media y tardía no refleja con exactitud la adecuación del CL.17,18
La respuesta de los tejidos esteroidogénicos a las hormonas gonadotrópicas está regulada en parte por los receptores en las células y tejidos gonadales. Como resultado de la unión de la hormona al receptor, la enzima adenilato ciclasa activa un aumento en los niveles intracelulares de AMPc. La actividad incrementada de la proteína cinasa conduce a una mayor síntesis de hormonas esteroides ováricas a través de varios mecanismos, incluyendo la activación directa de componentes de la enzima mediante la fosforilación. En el CL son varios los factores que influyen en el número de receptores a LH, entre ellos la activación directa de los componentes del sistema enzimático esteroidogénico vía fosforilación. El complejo hormona-receptor se internaliza entonces por endocitosis y la hormona se degrada en lisosomas. Después de la internalización, el receptor es también degradado en lisosomas o reciclado a través del aparato de Golgi. Los receptores nuevos o reciclados para LH se incorporan en la membrana limitante con proteínas que contienen gránulos secretores. Las acciones de la LH en la membrana plasmática de las células que contienen estos gránulos secretores hacen que aumente la exocitosis e incorporan estos gránulos en la membrana (y los receptores a LH). Estos mecanismos propuestos explican el aumento del número de receptores a LH inmediatamente después de la estimulación de la célula lútea con dosis masivas de LH. También explican la "regulación a la baja" de los receptores de LH, 24 horas después de la administración de LH.19
El conocimiento sobre los eventos moleculares que conducen a la regresión funcional y estructural del CL es limitado debido a que existen diferencias entre los eventos luteolíticos de mamíferos primates y no primates. El CL de los primates tiene una vida media de 14 -16 días en los ciclos menstruales no fértiles, pero en los ciclos menstruales en los que se produce una implantación, la continuación de su capacidad funcional es indispensable para el mantenimiento satisfactorio del embarazo, hasta que la placenta se convierta en la fuente principal de progesterona. Para aportar conocimiento sobre el mecanismo fisiológico por el cual la hCG prolonga la vida funcional del CL de primates, en monos Cynomolgus se administró hCG o LH ya sea a una tasa constante o en una tasa que aumenta exponencialmente durante la fase lútea media y tardía del ciclo menstrual. Los resultados indican que el CL experimenta una reducción sustancial en su capacidad de respuesta a la LH entre las fases lútea media y tardía del ciclo menstrual y que la capacidad de respuesta disminuida del CL es estas etapas solo puede superarse por las concentraciones elevadas de gonadotropinas logradas por la producción placentaria de la hCG.20,21
La regresión del CL está compuesta de dos eventos: la pérdida de su capacidad esteroidogénica (principalmente de progesterona), conocida como regresión funcional, seguida de una regresión estructural que involucra muerte celular por apoptosis y degeneración de la matriz extracelular por medio de luteólisis.22 La regresión del CL, tanto funcional como estructuralmente, es esencial para la ciclicidad normal. El CL debe retroceder funcionalmente para disminuir la secreción de progesterona y permitir otra oleada ovulatoria de LH y, por lo tanto, otro ciclo estral. También debe replegarse de manera estructural (destrucción y remoción de las células lúteas) para mantener el ovario en la proporción adecuada con respecto al resto del tracto reproductivo y habilitar los folículos preovulatorios para crecer y posteriormente ovular.23 Por lo anterior, podemos decir que el CL es una glándula endocrina cuya vida útil está programada por limitantes hormonales.24
La expresión del receptor CD95 (Apo-1) (Fas) y la apoptosis inducida por Fas son mecanismos atribuidos a la destrucción selectiva de células del CL durante la regresión lútea. En ciertos tipos de células epiteliales, la sensibilidad de estas se debe a la pérdida o escisión de los filamentos intermedios que contienen queratina. Específicamente la queratina 8/18 (K8/K18) de los filamentos influye en la muerte celular, en parte mediante la regulación de la expresión de Fas en la superficie celular.25 Sin embargo, no se han estudiado completamente los sistemas de señalización que impulsan la regresión del CL de los primates en los ciclos de no concepción.
Recientemente ha incrementado el interés por conocer el papel funcional de los metabolitos del estradiol (EM), principalmente en tejidos productores de estrógeno. El CL humano produce una serie amplia de EM, y se ha postulado que actúan a través de vías paracrinas y autocrinas que afectan la angiogénesis o los eventos mediados por LH.6,7
Por otra parte, se ha observado que el sistema inmune participa en la autofagositosis de células lúteas en humanos. Los macrófagos parecen jugar un papel indispensable en funciones ováricas, puesto que estos invaden al CL. La acumulación de células inmunes se asocia con la necrosis que se presenta durante la regresión del CL, como se ha observado en ovarios de vacas y humanos.26 Se sugiere que en el CL existe un proceso de necrosis ya que se producen grandes cantidades de especies reactivas de oxígeno (ROS); además, los altos niveles de cortisol causan estrés oxidativo en las células de la granulosa causando su muerte, lo que afecta directamente a los ovocitos ya que las células de la granulosa dejan de proveer a los folículos de nutrientes, factores de crecimiento y factores de supervivencia, lo que conlleva a la disminución en la producción de E2. Algunos estudios sugieren que el E2 participa como antioxidante en la supervivencia de los ovocitos de cerdo.27,28 La dependencia continua del CL de los primates sobre LH/ hCG /AMPc también parece estar detrás de la luteólisis, ya que parece requerir un mayor soporte luteotrópico.29
En contraste, la regresión del CL de los rumiantes es iniciada por prostaglandinas F-α (PGF) a partir de la señal del útero que no presenta implantación. En consecuencia, la fase lútea corta en rumiantes se debe principalmente a la secreción prematura de PGF por el útero no preñado, mientras que la insuficiencia del CL en primates está relacionada con un inadecuado soporte luteotrópico y a la regresión prematura del CL. Así, el CL tiene dos vías que dependen de la preñez: funcionar como el principal soporte de progesterona o la regresión del CL en la ausencia de la preñez, estos procesos son producidos por vías celulares y enzimáticas comunes, reguladas por procesos luteotrópicos y luteolíticos.30,31 Estos procesos incluyen factores paracrinos o autocrinos asociados con la angiogénesis, que median las acciones de LH/hCG (por ejemplo, progesterona), o contrarrestar efectos luteotrópicos (es decir, luteólisis local; por ejemplo, PGF2α).31
La inervación del ovario incluye componentes somáticos y autonómicos que llegan a los ovarios por medio del nervio ovárico superior y el nervio del plexo ovárico (NPO).32,33 Recientemente nuestro grupo de trabajo hizo una descripción detallada de la inervación extrínseca que llega a los ovarios a través del NPO, mostrando su diversidad en cuanto a los ganglios prevertebrales donde se localizan las neuronas posganglionares.34 Las terminales nerviosas localizadas en los ovarios liberan neurotransmisores y neuropéptidos que influyen en la actividad de los tejidos ováricos cuyo papel sería de tipo regulador en la esteroidogénesis y el desarrollo folicular.35-39 Así mismo se ha mostrado que en las membranas de las células foliculares existen receptores adrenérgicos y que, el empleo de fármacos agonistas o antagonistas modifica la liberación de los esteroides ováricos.40,41
La denervación del ovario disminuye la actividad esteroidogénica del CL, aumenta los receptores en las células lúteas, retrasa el desarrollo folicular e interrumpe la ciclicidad ovárica.37,39 Así mismo, la secreción de progesterona y de andrógenos se modifica cuando el ganglio celíaco es estimulado con agonistas noradrenérgicos como propanolol o fentolamina, o cuando se estimulan los receptores adrenérgicos en los ovarios.42 Incluso, se ha descrito que el efecto de la estimulación vía nerviosa supera al efecto producido por LH en la liberación de progesterona.43
Estudios in vitro e in vivo han mostrado que los receptores β-adrenérgicos modulan la liberación de hormonas esteroideas.44 En bovinos, el bloqueo de los β-receptores ováricos con propanolol disminuye las concentraciones de progesterona periférica entre el 20% al 30% a los 11 días de ser inyectado. Estos datos sugieren que la secreción basal de progesterona puede depender de la estimulación adrenérgica directa y constante en el CL.45
El sistema noradrenérgico puede participar en la función secretora del CL en varios niveles reguladores como pueden ser: (a) afectar el funcionamiento del CL a través de vías en el sistema nervioso central, que involucran áreas extrahipofisiarias, amigdalar e hipotálamo posterior; (b) controlar la contractilidad de la vasculatura periférica y ovárica, influyendo en el flujo sanguíneo de CL; y (c) aumentar la actividad de enzimas específicas que participan en la síntesis de oxitocina ovárica postraduccional y la esteroidogénesis del CL.13 En rumiantes, el funcionamiento del CL temprano es independiente de la LH, mientras que el CL en la fase lútea media requiere de LH como soporte luteotrófico, estos datos apuntan a la importancia de la estimulación β-adrenérgica en la regulación funcional del CL recién formado. Lo que supone que esto es especialmente importante en las primeras etapas de la preñez. Sin embargo, en novillas la infusión de un bloqueador β-adrenérgico a mitad del ciclo reduce la secreción de progesterona en un 20-30%. Por lo tanto, se ha sugerido que la estimulación β-adrenérgica es igualmente crucial para la función del CL durante toda la fase lútea como para todo el ciclo en general.13,45,46
8. Inervación intrínseca del ovario
La inervación intrínseca del ovario es provista por neuronas con soma multipolar alojadas principalmente en la médula ovárica; mientras que las localizadas en la corteza están inmersas en la capa de células de la teca, pero no atraviesan la membrana basal y por lo tanto, no ingresan al interior del folículo ni del CL.47
Respecto a la naturaleza bioquímica de las neuronas intrínsecas del ovario, se mostró que producen neuropéptido Y (NPY), aunque no se determinó si también producen otros péptidos.47 Resultados de nuestro laboratorio muestran que en los ovarios de la rata hembra adulta de la cepa Long Evans, existe una inervación intrínseca abundante y, específicamente en la corteza del ovario, cada folículo está asociado a un grupo de neuronas que pudiera influir en su desarrollo. También hemos encontrado que al interior del CL existen células que muestran inmunorreactividad a enzimas involucradas en la síntesis de catecolaminas (TH), que seguramente tienen la capacidad de influenciar la regulación y síntesis de progesterona en el CL.48
En el CL de mamíferos se ha sugerido que la dopamina (DA) puede tener influencia directamente a través de sus receptores específicos, indirectamente a través de los adrenoreceptores por medio de una reacción cruzada o mediante su conversión a NA en el proceso catalizada por la dopamina-β-hidroxilasa. La DA está presente en el CL de los bovinos, donde estimula la secreción de oxitocina en el CL de novillas.46 La DA estimula la liberación de oxitocina en la hipófisis de ratas lactantes, con una potencia comparable a la de la NA. De igual manera, se ha observado que la NA influye en la secreción concomitante de progesterona y oxitocina en el ovario durante todas las etapas del ciclo estral en bovinos actuando a través de los adrenoreceptores. De este modo suponemos que la DA afecta la función ovárica después de su conversión en NA.46,49
También se ha observado que la NA afecta la secreción de progesterona y su síntesis por mediante un aumento de la actividad del citocromo P450scc y de la 3β-hidroxiesteroide deshidrogenasa. Este efecto está mediado a través de los receptores celulares β1 y β2 de las células lúteas, la cantidad total de estos receptores se correlaciona con las concentraciones de progesterona periférica durante la fase lútea y esto refleja la capacidad del ovario para reaccionar a la estimulación.33,37,41,44
En otros estudios se ha observado que la denervación ovárica causa una disminución de la actividad esteroidogénica en el CL, un aumento de los receptores en las células lúteas, un retraso en el desarrollo folicular y la interrupción de la ciclicidad.37,39 El aumento de la actividad noradrenérgica durante situaciones estresantes podría afectar la función del CL y, por el contrario, el aumento duradero en las concentraciones de catecolaminas en sangre disminuye notablemente el número de receptores de catecolaminas en el CL, posiblemente por su autorregulación. Incluso se observó que las concentraciones de DA dentro del CL están altamente correlacionadas con los de NA durante el ciclo estral, y son más altas en el CL recién formado que en el CL desarrollado, el CL regresivo o el CL de mujeres embarazadas. El CL de bovinos puede sintetizar NA a partir de DA como precursor, también se ha observado que la respuesta del CL al tratamiento con NA se reduce en el envejecimiento, en comparación con las vacas cíclicas.45
La progesterona reduce la actividad de la MAO, la principal enzima responsable de la degradación intracelular de las catecolaminas.46 La MAO se encuentra alrededor de los vasos sanguíneos ováricos, en la glándula intersticial y en el CL en envejecimiento, pero no en los folículos o en el CL recién formado en ratas. La actividad de la MAO ovárica varía durante el ciclo estral en ratas,33 por lo que se supone que también puede afectar indirectamente la función ovárica.50 Lo que indica que la estimulación catecolaminérgica puede ser una parte importante en el mecanismo de secreción del CL.46,51
10. La insuficiencia de la Fase Lútea
La insuficiencia de la fase lútea debido a problemas hormonales lleva al fracaso de la implantación y ha sido responsable de abortos espontáneos y fracasos en reproducción asistida. Esta deficiencia se observa en mujeres con ovarios poliquísticos, trastornos de la tiroides y alteraciones en la producción de prolactina, lo cual genera un bajo ambiente hormonal de progesterona, provocando iatrogenia debido entre otros, a malas intervenciones en reproducción asistida. El uso de análogos de la hormona liberadora de gonadotropina para prevenir la oleada de LH y la aspiración de las células de la granulosa durante la recuperación del ovocito (en procedimientos de fertilización in vitro), puede perjudicar la capacidad del CL para producir progesterona. El uso de agentes progestacionales como progesterona/hCG han demostrado ser eficaces en mujeres con antecedentes de aborto recurrente. Sin embargo, no se ha demostrado un efecto beneficioso de agentes de uso generalizado para establecer un tratamiento óptimo, como ácido ascórbico, estrógeno o prednisolona junto con progesterona.14,21,52-54
Por otra parte, la obesidad y la malnutrición se han relacionado con una disminución de la fecundidad en las mujeres.55 La deficiencia de la capacidad reproductiva de mujeres obesas se atribuye a la anovulación. Sin embargo, en mujeres obesas con ciclos ovulatorios regulares se observa fertilidad reducida.56,57 Por lo cual, se sugiere que la obesidad afecta directamente la calidad de los ovocitos y embriones,58 así como la receptividad endometrial.59 También se ha visto que la excreción urinaria de metabolitos de progesterona disminuye, debido a la supresión en la secreción de gonadotropinas en mujeres obesas,60 por lo que se ha llegado a pensar que la obesidad afecta la función del CL. Además, se ha mostrado que la secreción de progesterona está asociada con la regulación negativa de las vías esteroidogénicas del CL, por lo que una variación de peso contribuye a la disfunción del CL.55 Este nuevo panorama nos ha ayudado a comprender mejor las consecuencias y factores asociados en la disfunción reproductiva relacionada con el peso, lo que podría ayudar a reforzar los tratamientos para mejorar la capacidad reproductiva de las mujeres con obesidad.53,54
En los últimos años se han realizado avances significativos sobre el estudio de la inervación y su influencia en diferentes órganos, incluyendo al ovario y sus estructuras, tales como el CL. La formación y duración del CL son críticas para la reproducción en mamíferos, ya que la progesterona lútea es esencial para la implantación del embrión y el mantenimiento de la preñez, hasta que se desarrolla la placenta. La disfunción de esta glándula dinámica puede generar problemas de salud tales como abortos por insuficiencia lútea, quistes ováricos e infertilidad.
Actualmente se han realizado avances significativos en el conocimiento de los mecanismos fisiológicos que regulan la función ovárica, sin embargo, se requieren más estudios que nos permitan conocer la neurobiología del CL, en especial para esclarecer las vías de señalización que subyacen para determinar la funcionalidad del CL en el ciclo ovárico. El conocimiento de la fisiopatología del CL requiere de estudios integrativos con enfoques metodológicos novedosos, in vivo e in vitro, que nos permitan conocer más de la insuficiencia lútea, los mecanismos celulares y endocrinos en el ovario que están siendo afectados por el estrés y que a su vez provocan patologías que antes tenían una menor prevalencia entre las mujeres en edad reproductiva. Resultados recientes y trabajos futuros pueden ser importantes no sólo para tratamiento de los trastornos ováricos en mujeres, sino que también para el diseño de nuevos anticonceptivos e incluso usarse para la recuperación de la fauna silvestre, especialmente en la reproducción de las especies en peligro de extinción. En la presente revisión se hizo énfasis en los mecanismos de regulación del CL y cómo participan en el control de las funciones ováricas como el ciclo ovárico y su influencia en posibles patologías asociadas al CL, dándonos una visión más amplia de lo que aún falta por estudiar para entender de los mecanismos de regulación del ovario.
Los autores declaran que no existen conflictos de interés.
Agradecemos al “Programa Nacional de Becas de Doctorado, CONACYT” por el apoyo para la realización de este trabajo CONACYT FC2016-01-231 y VIEP-3180. El autor Juan Manuel Bravo-Benítez fue apoyado con el programa de becas de doctorado de CONACYT-JMBB772933/624536.
1. Stouffer, R. L.; Bishop, C. V.; Bogan, R. L.; Xu, F.; Hennebold, J. D., Endocrine and local control of the primate corpus luteum. Reprod Biol 2013, 13 (4), 259-71.
2. Williams, C. J.; Erickson, G. F., Morphology and Physiology of the Ovary. In Endotext, Feingold, K. R.; Anawalt, B.; Boyce, A.; Chrousos, G.; Dungan, K.; Grossman, A.; Hershman, J. M.; Kaltsas, G.; Koch, C.; Kopp, P.; Korbonits, M.; McLachlan, R.; Morley, J. E.; New, M.; Perreault, L.; Purnell, J.; Rebar, R.; Singer, F.; Trence, D. L.; Vinik, A.; Wilson, D. P., Eds. South Dartmouth (MA), 2000.
3. Christenson, L. K.; Devoto, L., Cholesterol transport and steroidogenesis by the corpus luteum. Reprod Biol Endocrinol 2003, 1, 90.
4. Schams, D.; Berisha, B., Regulation of corpus luteum function in cattle--an overview. Reprod Domest Anim 2004, 39 (4), 241-51.
5. Bergers, G.; Benjamin, L. E., Tumorigenesis and the angiogenic switch. Nat Rev Cancer 2003, 3 (6), 401-10.
6. Reynolds, L. P.; Grazul-Bilska, A. T.; Redmer, D. A., Angiogenesis in the corpus luteum. Endocrine 2000, 12 (1), 1-9.
7. Robinson, R. S.; Woad, K. J.; Hammond, A. J.; Laird, M.; Hunter, M. G.; Mann, G. E., Angiogenesis and vascular function in the ovary. Reproduction 2009, 138 (6), 869-81.
8. Galvao, A. M.; Ferreira-Dias, G.; Skarzynski, D. J., Cytokines and angiogenesis in the corpus luteum. Mediators Inflamm 2013, 2013, 420186.
9. Singhal, S. S.; Singhal, J.; Yadav, S.; Dwivedi, S.; Boor, P. J.; Awasthi, Y. C.; Awasthi, S., Regression of lung and colon cancer xenografts by depleting or inhibiting RLIP76 (Ral-binding protein 1). Cancer Res 2007, 67 (9), 4382-9.
10. Awasthi, S.; Singhal, S. S.; Yadav, S.; Singhal, J.; Drake, K.; Nadkar, A.; Zajac, E.; Wickramarachchi, D.; Rowe, N.; Yacoub, A.; Boor, P.; Dwivedi, S.; Dent, P.; Jarman, W. E.; John, B.; Awasthi, Y. C., RLIP76 is a major determinant of radiation sensitivity. Cancer Res 2005, 65 (14), 6022-8.
11. Awasthi, S.; Singhal, S. S.; Singhal, J.; Yang, Y.; Zimniak, P.; Awasthi, Y. C., Role of RLIP76 in lung cancer doxorubicin resistance: III. Anti-RLIP76 antibodies trigger apoptosis in lung cancer cells and synergistically increase doxorubicin cytotoxicity. Int J Oncol 2003, 22 (4), 721-32.
12. Billhaq, D. H.; Lee, S., A potential function of RLIP76 in the ovarian corpus luteum. J Ovarian Res 2019, 12 (1), 34.
13. Kotwica, J.; Skarzynski, D.; Jaroszewski, J., Involvement of beta-adrenoceptors in the regulation of luteal function in cattle. Br Vet J 1991, 147 (3), 189-96.
14. Silva, J. F.; Ocarino, N. M.; Serakides, R., Luteal activity of pregnant rats with hypo-and hyperthyroidism. J Ovarian Res 2014, 7, 75.
15. Niswender, G. D.; Juengel, J. L.; Silva, P. J.; Rollyson, M. K.; McIntush, E. W., Mechanisms controlling the function and life span of the corpus luteum. Physiol Rev 2000, 80 (1), 1-29.
16. Clapp, C.; Thebault, S.; Macotela, Y.; Moreno-Carranza, B.; Triebel, J.; Martinez de la Escalera, G., Regulation of blood vessels by prolactin and vasoinhibins. Adv Exp Med Biol 2015, 846, 83-95.
17. Filicori, M.; Butler, J. P.; Crowley, W. F., Jr., Neuroendocrine regulation of the corpus luteum in the human. Evidence for pulsatile progesterone secretion. J Clin Invest 1984, 73 (6), 1638-47.
18. Yang, Y. L.; Ren, L. R.; Sun, L. F.; Huang, C.; Xiao, T. X.; Wang, B. B.; Chen, J.; Zabel, B. A.; Ren, P.; Zhang, J. V., The role of GPR1 signaling in mice corpus luteum. J Endocrinol 2016, 230 (1), 55-65.
19. Niswender, G. D., Response of the corpus luteum to luteinizing hormone. Environ Health Perspect 1981, 38, 47-50.
20. Zeleznik, A. J., In vivo responses of the primate corpus luteum to luteinizing hormone and chorionic gonadotropin. Proceedings of the National Academy of Sciences of the United States of America 1998, 95 (18), 11002-7.
21. Zeleznik, A. J., In vivo responses of the primate corpus luteum to luteinizing hormone and chorionic gonadotropin. Proc Natl Acad Sci U S A 1998, 95 (18), 11002-7.
22. Quirk, S. M.; Cowan, R. G.; Harman, R. M., Role of the cell cycle in regression of the corpus luteum. Reproduction 2013, 145 (2), 161-75.
23. Juengel, J. L.; Garverick, H. A.; Johnson, A. L.; Youngquist, R. S.; Smith, M. F., Apoptosis during luteal regression in cattle. Endocrinology 1993, 132 (1), 249-54.
24. Telleria, C. M., Can luteal regression be reversed? Reprod Biol Endocrinol 2006, 4, 53.
25. Duncan, A.; Forcina, J.; Birt, A.; Townson, D., Estrous cycle-dependent changes of Fas expression in the bovine corpus luteum: influence of keratin 8/18 intermediate filaments and cytokines. Reprod Biol Endocrinol 2012, 10, 90.
26. Bagnjuk, K.; Mayerhofer, A., Human Luteinized Granulosa Cells-A Cellular Model for the Human Corpus Luteum. Front Endocrinol (Lausanne) 2019, 10, 452.
27. Grootjans, S.; Vanden Berghe, T.; Vandenabeele, P., Initiation and execution mechanisms of necroptosis: an overview. Cell Death Differ 2017, 24 (7), 1184-1195.
28. Chaudhary, G. R.; Yadav, P. K.; Yadav, A. K.; Tiwari, M.; Gupta, A.; Sharma, A.; Pandey, A. N.; Pandey, A. K.; Chaube, S. K., Necroptosis in stressed ovary. J Biomed Sci 2019, 26 (1), 11.
29. Nichols, S. M.; Bavister, B. D.; Brenner, C. A.; Didier, P. J.; Harrison, R. M.; Kubisch, H. M., Ovarian senescence in the rhesus monkey (Macaca mulatta). Hum Reprod 2005, 20 (1), 79-83.
30. Wiltbank, M. C.; Salih, S. M.; Atli, M. O.; Luo, W.; Bormann, C. L.; Ottobre, J. S.; Vezina, C. M.; Mehta, V.; Diaz, F. J.; Tsai, S. J.; Sartori, R., Comparison of endocrine and cellular mechanisms regulating the corpus luteum of primates and ruminants. Anim Reprod 2012, 9 (3), 242-259.
31. Sansing, L. H.; Harris, T. H.; Kasner, S. E.; Hunter, C. A.; Kariko, K., Neutrophil depletion diminishes monocyte infiltration and improves functional outcome after experimental intracerebral hemorrhage. Acta Neurochir Suppl 2011, 111, 173-8.
32. Baljet, B.; Drukker, J., The extrinsic innervation of the abdominal organs in the female rat. Acta Anat (Basel) 1979, 104 (3), 243-67.
33. Aguado, L. I., Role of the central and peripheral nervous system in the ovarian function. Microsc Res Tech 2002, 59 (6), 462-73.
34. Pastelin, C. F.; Rosas, N. H.; Morales-Ledesma, L.; Linares, R.; Dominguez, R.; Moran, C., Anatomical organization and neural pathways of the ovarian plexus nerve in rats. J Ovarian Res 2017, 10 (1), 18.
35. Dissen, G. A.; Romero, C.; Paredes, A.; Ojeda, S. R., Neurotrophic control of ovarian development. Microsc Res Tech 2002, 59 (6), 509-15.
36. Hsueh, A. J.; Adashi, E. Y.; Jones, P. B.; Welsh, T. H., Jr., Hormonal regulation of the differentiation of cultured ovarian granulosa cells. Endocr Rev 1984, 5 (1), 76-127.
37. Mayerhofer, A.; Dissen, G. A.; Costa, M. E.; Ojeda, S. R., A role for neurotransmitters in early follicular development: induction of functional follicle-stimulating hormone receptors in newly formed follicles of the rat ovary. Endocrinology 1997, 138 (8), 3320-9.
38. Dominguez, R.; Cruz, M. E.; Chávez-Genaro, R., Differences in the Ovulatory Ability Between the Right and Left Ovary Are Related to Ovarian Innervation. Growth Factors and the Ovary 1988, 39, 321-325.
39. Dominguez, R.; Riboni, L., Failure of ovulation in autografted ovary of hemispayed rat. Neuroendocrinology 1971, 7 (3), 164-70.
40. Gerendai, I.; Csaba, Z.; Vokó, Z.; Csernus, V., Involvement of a direct neural mechanism in the control of gonadal functions. The Journal of Steroid Biochemistry and Molecular Biology 1995, 53 (1), 299-305.
41. Ojeda, S. R.; Lara, H. E. In Role of the Sympathetic Nervous System in the Regulation of Ovarian Function, Berlin, Heidelberg, Springer Berlin Heidelberg: Berlin, Heidelberg, 1989; pp 26-32.
42. Casais, M.; Sosa, Z. Y.; Rastrilla, A. M.; Aguado, L. I., Coeliac ganglion adrenergic activity modifies ovarian progesterone during pregnancy: its inter-relationship with LH. J Endocrinol 2001, 170 (3), 575-84.
43. De Bortoli, M. A.; Garraza, M. H.; Aguado, L. I., Adrenergic intracerebroventricular stimulation affects progesterone concentration in the ovarian vein of the rat: participation of the superior ovarian nerve. J Endocrinol 1998, 159 (1), 61-8.
44. Aguado, L. I.; Petrovic, S. L.; Ojeda, S. R., Ovarian β-Adrenergic Receptors during the Onset of Puberty: Characterization, Distribution, and Coupling to Steroidogenic Responses*. Endocrinology 1982, 110 (4), 1124-1132.
45. Kotwica, J.; Bogacki, M.; Rekawiecki, R., Neural regulation of the bovine corpus luteum. Domest Anim Endocrinol 2002, 23 (1-2), 299-308.
46. Kotwica, J.; Skarzynski, D.; Bogacki, M.; Miszkiel, G., Influence of dopamine as noradrenaline precursor on the secretory function of the bovine corpus luteum in vitro. Br J Pharmacol 1996, 118 (7), 1669-74.
47. D'Albora, H.; Anesetti, G.; Lombide, P.; Dees, W. L.; Ojeda, S. R., Intrinsic neurons in the mammalian ovary. Microsc Res Tech 2002, 59 (6), 484-9.
48. Nayak, D.; Roth, T. L.; McGavern, D. B., Microglia development and function. Annu Rev Immunol 2014, 32, 367-402.
49. Tripathy, S.; Asaithambi, K.; Jayaram, P.; Medhamurthy, R., Analysis of 17beta-estradiol (E2) role in the regulation of corpus luteum function in pregnant rats: Involvement of IGFBP5 in the E2-mediated actions. Reprod Biol Endocrinol 2016, 14, 19.
50. Yoshimoto, Y.; Sakumoto, T.; Arai, R.; Miyake, A.; Kimura, H.; Aono, T.; Tanizawa, O.; Maeda, T., Monoamine oxidase in rat ovary during the estrous cycle. A histochemical study by a new coupled peroxidatic oxidation method. Endocrinology 1986, 119 (4), 1800-4.
51. Dumesic, D. A.; Richards, J. S., Ontogeny of the ovary in polycystic ovary syndrome. Fertil Steril 2013, 100 (1), 23-38.
52. Ojeda, S. R.; Lara, H. E. In Role of the Sympathetic Nervous System in the Regulation of Ovarian Function, The Menstrual Cycle and Its Disorders, Berlin, Heidelberg, 1989//; Pirke, K. M.; Wuttke, W.; Schweiger, U., Eds. Springer Berlin Heidelberg: Berlin, Heidelberg, 1989; pp 26-32.
53. Lustig, R. H.; Bradlow, H. L.; Fishman, J. In Estrogen Metabolism in Disorders of Nutrition and Dietary Composition, The Menstrual Cycle and Its Disorders, Berlin, Heidelberg, 1989//; Pirke, K. M.; Wuttke, W.; Schweiger, U., Eds. Springer Berlin Heidelberg: Berlin, Heidelberg, 1989; pp 119-132.
54. Shah, D.; Nagarajan, N., Luteal insufficiency in first trimester. Indian J Endocrinol Metab 2013, 17 (1), 44-9.
55. Kuokkanen, S.; Polotsky, A. J.; Chosich, J.; Bradford, A. P.; Jasinska, A.; Phang, T.; Santoro, N.; Appt, S. E., Corpus luteum as a novel target of weight changes that contribute to impaired female reproductive physiology and function. Syst Biol Reprod Med 2016, 62 (4), 227-42.
56. Polotsky, A. J.; Hailpern, S. M.; Skurnick, J. H.; Lo, J. C.; Sternfeld, B.; Santoro, N., Association of adolescent obesity and lifetime nulliparity--the Study of Women's Health Across the Nation (SWAN). Fertil Steril 2010, 93 (6), 2004-11.
57. Bolumar, F.; Olsen, J.; Rebagliato, M.; Saez-Lloret, I.; Bisanti, L., Body mass index and delayed conception: a European Multicenter Study on Infertility and Subfecundity. Am J Epidemiol 2000, 151 (11), 1072-9.
58. Bellver, J.; Pellicer, A.; Garcia-Velasco, J. A.; Ballesteros, A.; Remohi, J.; Meseguer, M., Obesity reduces uterine receptivity: clinical experience from 9,587 first cycles of ovum donation with normal weight donors. Fertil Steril 2013, 100 (4), 1050-8.
59. Bellver, J.; Martinez-Conejero, J. A.; Labarta, E.; Alama, P.; Melo, M. A.; Remohi, J.; Pellicer, A.; Horcajadas, J. A., Endometrial gene expression in the window of implantation is altered in obese women especially in association with polycystic ovary syndrome. Fertil Steril 2011, 95 (7), 2335-41, 2341 e1-8.
60. Santoro, N.; Lasley, B.; McConnell, D.; Allsworth, J.; Crawford, S.; Gold, E. B.; Finkelstein, J. S.; Greendale, G. A.; Kelsey, J.; Korenman, S.; Luborsky, J. L.; Matthews, K.; Midgley, R.; Powell, L.; Sabatine, J.; Schocken, M.; Sowers, M. F.; Weiss, G., Body size and ethnicity are associated with menstrual cycle alterations in women in the early menopausal transition: The Study of Women's Health across the Nation (SWAN) Daily Hormone Study. J Clin Endocrinol Metab 2004, 89 (6), 2622-31.
| Recibido: 26 de julio de 2021 | Aceptado: 17 de octubre de 2021 |
*Correspondencia: Morán Raya Carolina. Laboratorio de Aplicaciones Biomédicas, Centro de Investigación en Fisicoquímica de Materiales, Instituto de Ciencias, Benemérita Universidad Autónoma de Puebla, Puebla, México. Dirección: 72570, Av. San Claudio 1814, Jardines de San Manuel, 72570 Puebla, Pue. Teléfono: 2223611400 E-mail: carolina.moran@correo.buap.mx
Este es un artículo de libre acceso distribuido bajo los términos de la licencia de Creative Commons, (http://creamasal@unam.mxtivecommons.org/licenses/by-nc/3.0), que permite el uso no comercial, distribución y reproducción en algún medio, siempre que la obra original sea debidamente citada.