Artificial vagino-cervical stimulation modifies the frequency of uterine contractions during the estrous cycle of the female rat
- Inicio
- Comité Editorial
- Lineamientos
- Carta de Cesión de Derechos
- Información Legal
- Acerca de la Revista
- Bases de Datos
- Contacto
- ISSN 2007-3054
- Centro de Investigaciones Cerebrales
Universidad Veracruzana
Carrasco-García AA1, Camacho MA2, Pacheco P2, Carrillo P2, Coria-Avila GA3, *Paredes-Ramos P1
1Facultad de Medicina Veterinaria y Zootecnia, Universidad Veracruzana, Veracruz, Veracruz, México. 2Instituto de Neuroetología, Universidad Veracruzana, Xalapa, Veracruz. México. 3Centro de Investigaciones Cerebrales, Universidad Veracruzana, Xalapa, Veracruz, México
Resumen/Abstract
Introduction
Methods
Statistical analysis
Results
Discussion
Conclusions
References
Correspondencia
Las contracciones uterinas (CU) pueden participar en importantes procesos reproductivos, como el transporte de espermatozoides y el parto. La evidencia indica que las CU pueden ser inducidas por hormonas y estimulación vaginocervical (EVC) a través de vías neuroendocrinas, incluida la actividad de las glándulas pituitaria y ovarios. Evaluamos la frecuencia de CU después de la EVC en diferentes fases del ciclo estral (CE) en ratas hembras adultas. Treinta y seis ratas hembras fueron anestesiadas, y sus cuernos uterinos fueron expuestos a través de una incisión abdominal. La EVC incluyó intromisiones vaginocervicales (IVC) y distensión vaginal mediante la inserción de un tapón de silicón para imitar los efectos del tapón posteyaculatorio. El número de CU se midió: 1) antes y después de las IVC, 2) durante la distensión vaginal por el tapón de silicón, y después de que se retiró. Comparamos el efecto de la EVC en la cantidad de CU durante las diferentes etapas de la CE de ratas. Los resultados indicaron que la frecuencia de la CU basal fue mayor durante el proestro tardío y menor durante el diestro. Además, las intromisiones dieron como resultado una mayor actividad uterina. La EVC aumentó el número de contracciones durante todas las fases de la CE, excepto durante el diestro. Discutimos el papel de las hormonas y la estimulación sexual en la CU, y su posible implicación en los procesos reproductivos de las hembras.
Palabras clave: Estimulación vaginocervical, Contracciones uterinas, Ciclo estral, Rata hembra.
Uterine contractions (UC) can participate in important reproductive processes such as sperm transport and parturition. Evidence indicates that UC can be induced by hormones and vaginocervical-stimulation (VCS) via neuroendocrine pathways, including the activity of the pituitary and reproductive glands. The frequency of UC was evaluated after VCS at different phases of the estrous cycle (EC) in adult female rats. Thirty-six female rats were anesthetized, and the uterine horns were exposed via abdominal incision. VCS included vaginocervical intromissions (VCI) and vaginal distention via insertion of a silicon plug to mimic the effects of the post-ejaculatory sperm plug. The number of UC was measured 1) before and after VCI, 2) during the vaginal distention by the silicon plug and after it was removed. The effect of VCS on the amount of UC during different stages of the rat EC was compared. Results indicated that the frequency of baseline UC was higher during late-proestrus and lower during diestrus. In addition, intromissions resulted in higher uterine activity. Interestingly, VCS increased the number of contractions during all phases of the EC, except during diestrus. The role of hormones and sexual stimulation on UC, and the potential implication in reproductive processes in females are discussed.
Keywords: Vaginocervical stimulation, Uterine contractions, Estrous cycle, Female rat.
1. Introduction
During sexual behavior, female rats receive vaginocervical stimulation (VCS) by intromissions from the male and the sperm plug formed after ejaculation. Stimulation by intromissions requires pelvic thrusts in order to penetrate the vagina with the penis. It consists of fast penetrations, approximately 0.3 seconds in duration and time intervals of 0.5 to 1 minute. Male rats perform from 8 to 15 intromissions before ejaculation.1,2 After that, seminal fluid coagulates in the reproductive female tract and forms a sperm plug that distends the vagina for a short period. It has been observed that both intromissions and the sperm plug are able to increase the number of uterine contractions (UC) in rats.3
Sperm plug weights around 115 mg has a conic shape with a height of 12.41 ± 0.13 mm and 5.31 ± 0.05 mm wide.4,5 Extirpation of the seminal vesicles and coagulating glands, prevent plug formation, and has deleterious effects on fertility and fecundity.6 According to studies6,7 sperm plug formation is involved in different functions including the gradual release of spermatozoa within the female tract, prevention of semen leakage and stoppage of subsequent insemination by other males.
It has been shown that the amount and pattern of VCS received during copulation can induce diverse behavioral and neuroendocrine activity in female rats. For instance, while few intromissions can reduce the duration of the estrous cycle (EC),8 facilitate lordosis,9 and induce analgesia,10,11 moderate amounts of VCS improve transcervical sperm transport,12-14 and increase the release of luteinizing hormones, oxytocin and prolactin in the serum of female rat.15-17 In addition, the number of intromissions received during copulation can be crucial for successful reproduction in different species. It has been demonstrated that the induction of pregnancy or pseudopregnancy in the golden hamster is related to the number and rate of intromissions received by the females.18 Moreover, different combinations of intromission frequency, as well as the duration of each insertion, and interval between insertions, lead to different percentages of success to induce pseudopregnancy in mice.19 In the female rat, the number of intromissions received during copulation can shorten the duration of sexual receptivity 20 and facilitates the implantation of the blastocyst in the uterus.13 Altogether, it indicates that the behavioral and neuroendocrine activity triggered as consequence of sexual stimulation depends on the amount and pattern of VCS received during copulation.
On the other hand, sperm transport is a critical step for reproduction. To fertilize the ovules, sperms cells need to overcome multiple obstacles including acidic pH, immune responses and oxidative damage in the reproductive female tract.12-14 Evidence indicates that VCS can facilitate the transport of sperm toward the oviducts by increasing uterine mobility.21 It has been showed that copulation induces a high frequency of strong peristaltic and non-peristaltic circular myometrial contractions propagating both cranially and caudally, which facilitate sperm transport and fertilization.22 In addition, artificial VCS increase the number of peristalsis of the uterine tract, but of a lower intensity as compared to the response elicited by a typical intromission performed by a male rat during copulation.22
Evidence indicates that uterine activity can be affected by several factors including age and hormonal status of the female.12-141517,23 Using a nonsurgical catheter technique for measurement of the uterine intraluminal pressure changes, it has been observed24 that multiparous cyclic sows progressively increased and decreased the amount of UC according to the phase of the estrous cycle (EC). Data showed that from a few days before estrus until estrus, the percentage of females showing uterine activity increased from 55 to 100%. Furthermore, the frequency and amplitude of UC for sows increased from 15 to 22/h, and from 20 to 40 mmHg on average. This pattern of uterine activity increased from proestrus to estrus and declined during the first days after estrus. Finally, the authors observed that intrauterine applications of hormones, such as estrogen and cloprostenol, a synthetic analogue of prostaglandin F2α, increased uterine activity in the sows. The study concluded that spontaneous UC increase around estrus and sex hormones can directly regulate uterine mobility during the EC in this species.
Despite the evident importance of uterine mobility, a limited number of studies have evaluated the role of sexual stimulation on the activity of the female reproductive tract. The present study was designed to determine the modulatory effects of artificial VCS on the frequency of UC, during different stages of the EC in rats.
2.1 Animals
Thirty sexually naive female Wistar rats (250-300g) were locally bred and housed at the Instituto de Neuroetología, Universidad Veracruzana, in groups of three to five per cage (50x30x20 cm) containing wood chip bedding and kept in a room maintained at 24ºC ± 2ºC, on a reversed 12 h light/dark cycle (lights off at 08:00). Rodent chow 18% protein (Harlan, Mexico) and drinking water were available ad libitum. The experimental protocols in this study were approved by a committee of the graduate program in Neuroetología, Universidad Veracruzana Mexico, following the Official Mexican Norm NOM-062-ZOO-1999 (Technical Specifications for the Production, Care and Use of Laboratory Animals).
2.2. Estrus cycle determination
To identify the phases of the EC, vaginal smears were analyzed every day during the dark phase, two hours after the lights went off (10:00 h). Vaginal smears consisted in obtain drops with vaginal cells. The rat was held by the tail to expose the perianal area and 10 microliters of saline solution (0.9% NaCl) was dropped into the vagina three times. The last time the drops were recovered from the vagina and they were placed on a microscope slide and observed with a optical microscope at 10X. In this way we observed the type of cells and determined the stage of the estrous cycle of every rat. EC was divided in early-proestrus, late-proestrus, early-estrus, late-estrus (metaestrus) and diestrus, according to the characteristics and proportion of epithelial cells, cornified cells and leukocytes, as follows: Early-Proestrus, consisted in numerous leukocytes and flattened epithelial cells but few cornified cells. During late-proestrus cells were mostly nucleated epithelial cells and few cornified cells. Early-estrus was considered when most of the cells were cornified with few nucleated epithelial cells.25,26 Late-estrus consisted of numerous detachments of the cornified cells and few leukocytes. Finally, during diestrus most common cells were leukocytes, with few cornified cells and basophilic cells.26-28 We used this division considering that the sexual receptivity of the female rat occurs during late-proestrus, but not during early-proestrus.
2.3. Surgery
During the third EC, female rats were anesthetized to receive artificial VCS and register the number of UC. Anesthesia consisted of a mixture of ketamine hydrochloride (50 mg/ml) and Xylazine hydrochloride (4 mg/ml), injected intraperitoneally in a volume of 1 ml/kg of body weight. Rats were placed in the supine position and received a midline incision from the xiphoid process to the pubic symphysis. Intestines were displaced outside of the abdominal cavity and kept warm and hydrated with a wet gauze containing a phosphate-buffered saline solution (37ºC). Likewise, the uterus was continuously dampened. Uterine contractions were recorded by laparoscopy with a Sony TRV608Hi8 Handycam digital videocamera using a 7x (24 squares/sec.) zoom and analyzed later.
2.4. Artificial vaginocervical stimulation
To generate VCS, we provided a pattern of artificial manipulation that attempted to mimic the intromissions and vaginal distention by the sperm plug provided by a male rat to the female during copulation. VCS included intromissions and the placement of a plug that distended the vagina. Intromissions were provided by a 4.5mm x 9.5cm glass rod lubricated with mineral oil introduced 22 mm into the vaginal orifice of the female. Stimulation by intromissions consisted of five 1-second insertions of the glass rod, alternating with 15 seconds of rest, for a total duration of 1 minute (see Figure 1). Ten minutes later a silicon plug (4.5 mm diameter, 12 cm height) was introduced into the vagina of the rats with a tooth stick and remained there by five minutes. The frequency of UC was determined in female rats instrumented during different phases of the EC. As illustrated in Figure 1, frequency of UC was measured during different epochs: 1) before intromissions (baseline), 2) after intromissions, 3) while the silicon plug was placed in the vagina, and 4) after the plug was removed.
Factorial effect analysis of Variance (ANOVA) was used to determine differences in the number of UC during the different patterns of VCS (Factor A), during the phases of the EC (factor B), and in the interaction between VCS and EC (A x B). For all significant main effects, Tukey HSD posthoc tests were conducted to assess differences between individual means. The level of significance for all comparisons was p<0.05.
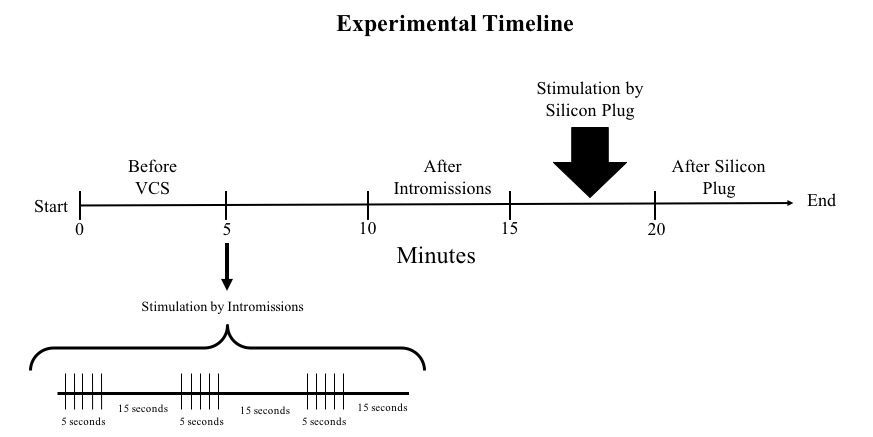
Figure 1. Experimental timeline. The number of uterine contractions (UC) was compared during the following periods: Five minutes previous to stimulation by intromissions (Before VCS), five minutes after intromissions, five minutes while the silicon plug was placed into the vagina, and the following five minutes after the silicon plug was removed. Stimulation by intromissions consisted of five 1-second insertions, alternating with 15 seconds of rest, for 1 minute.
The ANOVA detected significant differences in the frequency of UC during the stages of the EC F (4, 32) = 47.9, p <0.00005. The posthoc test indicated that the highest number of UC occurred during late-proestrus, as compared with the rest of the phases. Moreover, during early-estrus females showed more UC than during late-estrus and diestrus. However, no differences between early-proestrus and late-estrus were observed. Finally, the lowest level of uterine activity was observed during the diestrus (Figure 2).
On the other hand significant differences in the number of UC during VCS were observed F (3, 32) = 4.59, p <0.005. The posthoc test revealed that before VCS less UC were observed as compared with after stimulation. Also, the highest number of UC was consistently observed after intromissions (Figure 3).
Finally, the statistical analysis detected a significant effect on the interaction of the VCS and the phases of the EC on the amount of UC F (12, 100) = 2.16, p < 0.05. Posthoc test revealed that during early proestrus the number of UC increased after intromissions compared with the rest of periods. Besides after intromissions and during plug placement the number of UC increased during late-proestrus, early estrus and late-estrus, but failed to increase during diestrus. Finally, the highest increment of UC was observed after intromissions during late-proestrus as compared with the rest of the interactions (table 1).
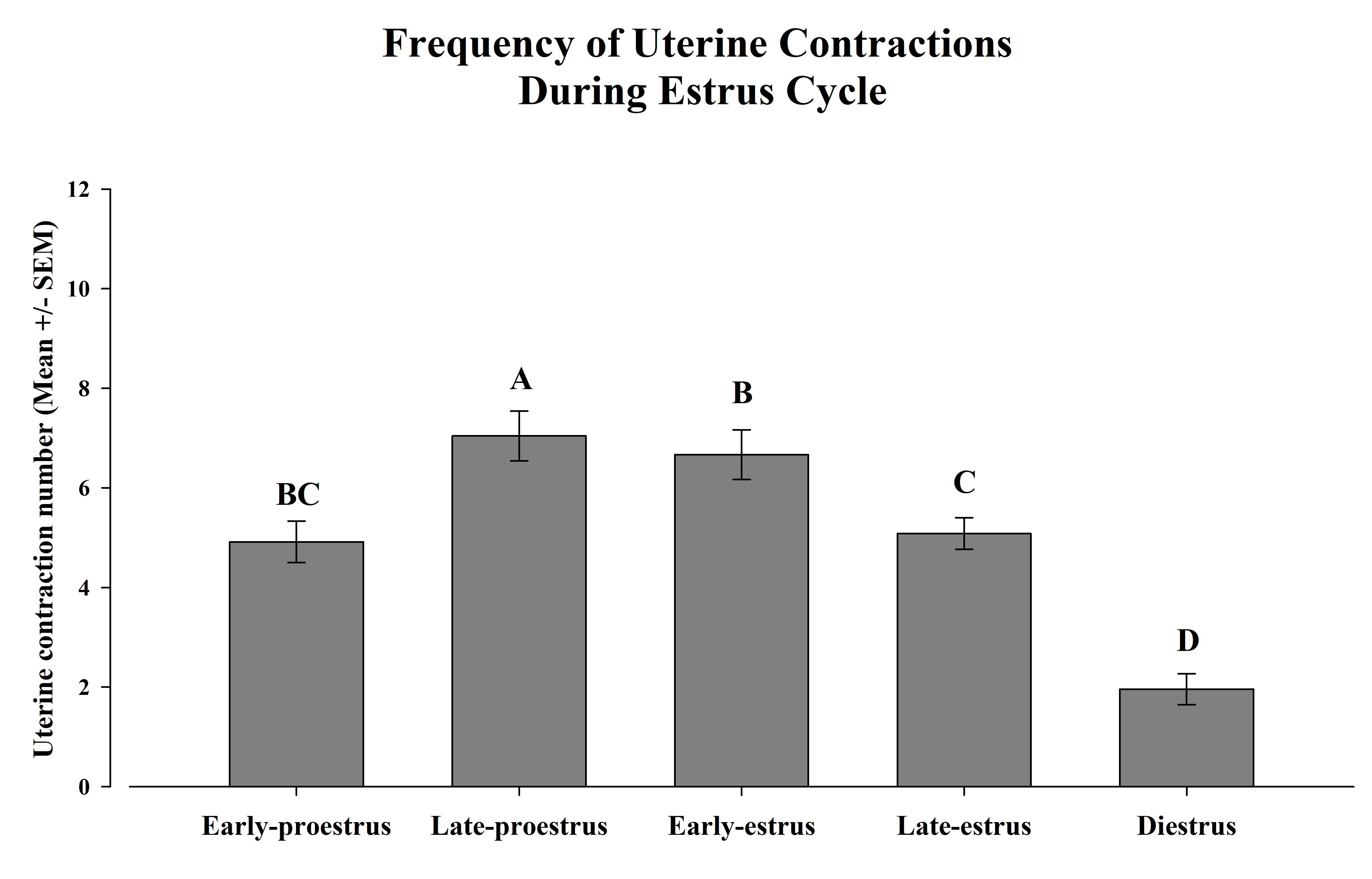
Figure 2. Frequency of uterine contractions during the estrus cycle. Variations in the frequency of uterine contractions during a different period of the estrus cycle in female rats. The frequency of uterine contractions (UC) in anesthetized female rats was determined by videography. The bars represent the mean number ± SEM of UC during the different phases of the EC. Different capital letters between bars indicate p<0.05.
The present study was designed to improve our understanding about the interactions between sexual stimulation and the EC on the uterine activity of female rats. Accordingly, our results demonstrated that VCS increased the frequency of UC during most phases of the EC, except at diestrus. Since the diestrus is characterized by a low hormonal activity, it clearly suggests that the effect of the VCS on the uterine activity depends on the levels and activity of hormones associated to EC in the female rat. On the other hand, the highest increment in the amount of UC as consequence of intromissions was observed during late-proestrus. Copulatory behavior is observed only during late-proestrus in rats. Consequently, we consider that the particular increment in the uterine activity during this period is oriented to facilitate reproductive processes such as sperm transport and fertilization. The role of hormones and sexual stimulation on the uterine activity, as well as the potential implication in reproductive processes is discussed as follows.
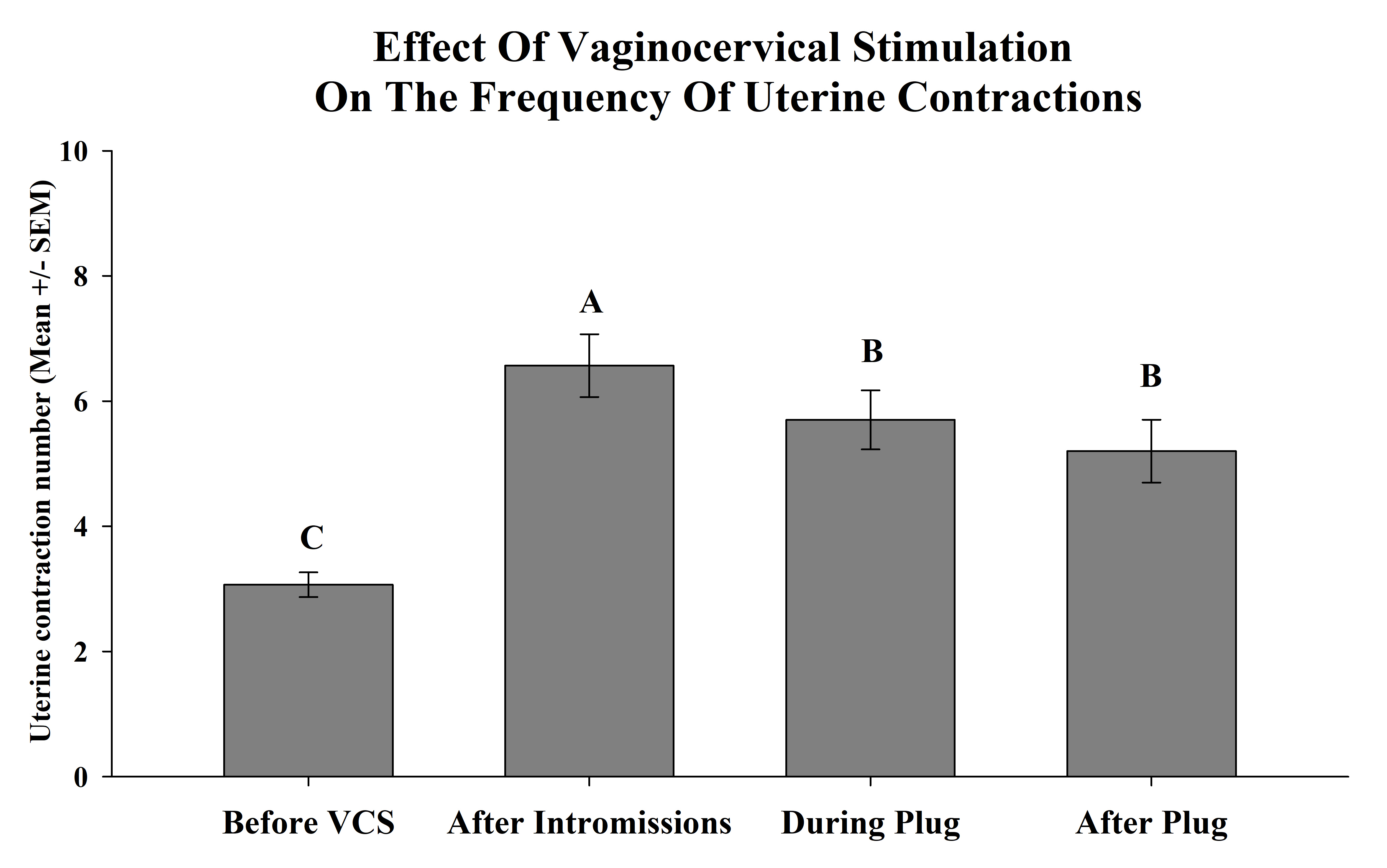
Figure 2. The frequency of UC was determined before vaginocervical stimulation (VCS), after intromissions, during the time the silicon plug was placed into the vagina and after it was removed. VCS increased the amount of UC, but fewer contractions were observed during plug placement, and after plug removal. The bars represent the mean number ± SEM of UC during 5-minute periods. Different capital letters between bars indicate p<0.05.
Hormones and Estrous Cycle
EC is defined as the time from the onset of one period of sexual receptivity to the next and includes changes in the activity of ovaries29 and brain areas related to sexual motivation.30 EC is mainly orchestrated by the action of ovarian hormones regulated by gonadotropins from the anterior pituitary.29,31,32 During proestrus, FSH secretion stimulates maturation of the follicles and both estrogen and LH levels increase as follicles mature. Estrogen released by ovarian follicles also increases the production of LH in the pituitary gland that during estrus stimulates ovulation and the ovarian follicles ruptures. Whereas estradiol decreases at late-proestrus, progesterone, FSH and LH remain elevated until late-estrus. Contrary to the rest of the stages of the EC, diestrus is characterized by a significant reduction in the levels of sexual hormones and by the lack of sexual motivation in females.29,32,33 On the other hand, evidence indicates an interesting interaction between non-ovarian hormones and the EC on the uterine activity of different animals. For instance, in a study, drug-induced uterine activity was recorded via uterine-implanted catheter-tip pressure transducers in bitches during proestrus and estrus, as well as during early and late diestrus. UC was registered in unanesthetized bitches during the EC. Results indicated that uterine activity was greatest during estrus, moderate during proestrus, and low during early and late diestrus. While intravenous injections of prostaglandin F2 alpha (5 micrograms/kg of body weight) induced intrauterine pressures of 107 mm of Hg during proestrus and 115 during estrus. Intravenous injections of oxytocin (0.05 USP units/kg,) induced pressures of 106 during proestrus and 116 mm of Hg during estrus. In contrast, the intrauterine pressure values induced by same hormones during early and late diestrus were 61 and 58 mm of Hg, respectively, and for oxytocin, were 61 and 51 mm of Hg, respectively. Authors concluded that in the bitch, drug-induced uterine activity are decreased during early and late diestrus.34 In addition, it has been observed that oxytocin levels change in serum during the estrous cycle, for example in the female sheep plasma concentrations of oxytocin increase during the luteal phase (proestrus) but decreased during estrus.35 Accordingly, our results indicated that the frequency of baseline UC increased during late-proestrus, decreasing progressively until diestrus. We believe the fluctuations in the levels of ovarian and non-ovarian hormones from proestrus to estrus facilitate the uterine activity and modify the threshold of reflex contractions when the female rat receives sexual stimulation. Since VCS failed in inducing UC during diestrus, it indicates that the increment in the amount of uterine activity as consequence of artificial VCS depends on the action and levels of hormones in the female rat. Nevertheless, future studies must be aimed at exploring the role of every single hormone on the effect of VCS on the uterine activity of females.
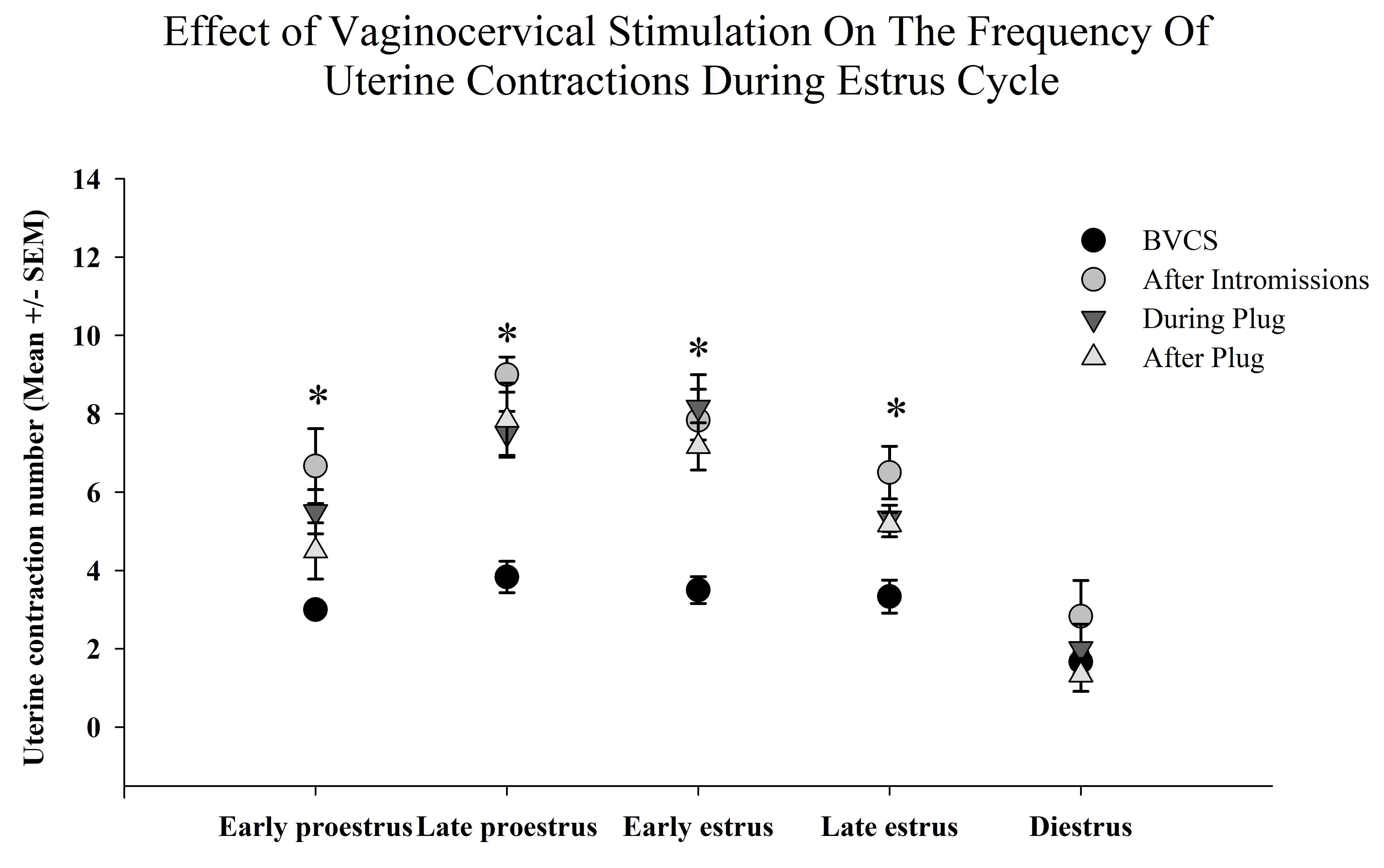
Table 1. Effect of vaginocervical stimulation on the frequency of uterine contractions during the estrus cycle. After intromissions and during plug the number of uterine contractions (UC) increased compared with the periods before vaginocervical stimulation (VCS) and after plug the different phases of the EC, except at diestrus. The highest increment of UC was observed after intromissions during late-proestrus as compared with the rest of the EC. Table shows the mean number ± SEM of UC by the different patterns of artificial VCS during the stages of the EC. Different capital letters indicates significant differences in the interaction of VCS and the phases of the EC p<0.05.
Hormones and female sexual behavior
Female rat sexual behavior includes proceptivity and receptivity.36 While proceptivity refers to behaviors aimed to bring the attention of the male rat (e.g. solicitation, hopping and darting), receptivity relates to the lordosis reflex (arching of the back, elevation of the rump, dorsiflexion of the tail, and extension of the neck) that is a stereotyped posture of the female, required for successful copulation.36 In the rat, both receptivity and proceptivity are observed during late-proestrus. Evidence indicates that estrogen and progesterone initiate sexual receptivity in the female rat. While estrogens allow the female to express lordosis, progesterone can intensify the expression of this behavior.31,36 However, if the amount of estrogen is sufficiently large in the rat, progesterone is not required for lordosis to occur.37 Accordingly, in our study, the highest increment in the number of UC as consequence of VCS occurred during late-proestrus. Thus, we believe that estrogen and progesterone are responsible for the facilitation of the effects of VCS on UC during late-proestrus. In addition, since rat sexual behavior occurs during late-proestrus, we believe the increment in the uterine activity could be oriented to facilitate reproductive processes such as transport of sperm, blastocysts and embryo implantation. The possible role and interaction of progesterone and estrogen on the uterine activity of female rats, though, still need to be examined in closer detail in future studies.
Effect of vaginocervical stimulation on reproductive processes
During copulation, sexual stimulation can trigger cascades of neuroendocrine activity8,10,12,15,17,30 that participate in different reproductive processes in animals.12,18,38,39 It has been observed that the induction of pregnancy or pseudopregnancy in the female rat depends on the number of intromissions received by the female during copulation.12 In addition, different combinations in the number of intromissions, the duration and the interval between insertions produce different percentages of pseudopregnancy in mice.19 Likewise, evidence indicates that the vaginal distention produced by the sperm plug after copula, extend the VCS beyond the distension caused by ejaculation alone, and it seems to affect reproductive processes such as sperm transport, sperm competition and fertilization.3,6,7,24 Vaginal distension activates a vagino-uterine reflex that induces UC, but it disappears if such distension is achieved for too long.40 A similar reflex in rats that aids sperm transport from the vagina to the uterus has been proposed.3 In our study, more uterine activity was observed during the periods related to vaginal distention by a silicon plug as compared with the baseline. However, vaginal distention decreased the amount of UC as compared with that observed after intromissions. Consequently, we believe that VCS in the female rat occurred for too long which reduced the capacity of the vagino-uterine reflex to induce UC. Nevertheless, we must consider that in our study the silicon plug was placed ten minutes after intromissions. Thus, it is not possible to determine if the reduction in the UC observed during the periods related to the plug (during plug and after plug) are consequence of vaginal distension achieved for too long, or if it is consequence of the elapsed time. In the future, different animals should be used to compare the effect of intromissions and vaginal distention on the UC of the rats.
The results observed in our experiment indicate that both VCS and hormonal status are important regulators of uterine mobility in rats. Since VCS was unable to increase UC during diestrus, we conclude that the effect of sexual stimulation on uterine activity strongly depends on the hormonal status of the female rat. Thus, the highest VCS-induced increment in UC occurred during late-proestrus. We suggest that myometrial activity is more responsive to sexual stimulation during the period where copula and reproductive processes can occur. Finally, we conclude that the uterine functionality of the female rat depends on both endogenous and exogenous stimuli. Future studies should evaluate the effects of VCS on reproductive processes such as sperm transport and fertilization, as well as on the release of gonadotropin and ovarian hormones.
This research was supported by studentship from CONACyT of Mexico to AACG. The author would like to thank Dr. Joseph Mauban for useful discussion and English grammar review.
The authors declare no conflict of interest. All authors have approved the final article and contributed to the research and/or article preparation.
1. Holmes LG, Himle MB, Sewell KK, Carbone PS, Strassberg DS, Murphy NA. Addressing Sexuality in Youth with Autism Spectrum Disorders: Current Pediatric Practices and Barriers. J Dev Behav Pediatrics. 2014;35(3):172.
2. Knickmeyer RC, Wheelwright S, Hoekstra R, Baron-Cohen S. Age of menarche in females with autism spectrum conditions. Dev Med Child Neurol. 2006;48(12):1007-1008.
3. Tordjman S, Anderson GM, McBride AP, Hertzig ME, Snow ME, Hall LM, Ferrari P, Cohen DJ. Plasma androgens in autism. J Autism Dev Disord. 1995;25(3):295-304.
4. Coleman M, Gillberg C. The Autisms. Oxford University Press; 2012.
5. Manzo J, Carrillo P, Coria-Avila GA, Garcia LI. The sexual cerebellum. En: Komisaruk BR and G Gonzalez-Mariscal, Behavioral neuroendocrinology. CRC Press 2017:103-112.
6. Hernandez ME, Soto-Cid A, Aranda-Abreu GE, Diaz R, Rojas F, Garcia LI, Toledo R, Manzo J. A study of the prostate, androgens and sexual activity of male rats. Reprod Biol Endocrin. 2007;5(1):11.
7. Manzo J, Vazquez MI, Cruz MR, Hernandez ME, Carrillo P, Pacheco P. Fertility ratio in male rats Effects after denervation of two pelvic floor muscles. Physiology Behav. 2000;68(5):611-618.
8. Rodier PM, Ingram JL, Tisdale B, Nelson S, Romano J. Embryological origin for autism: Developmental anomalies of the cranial nerve motor nuclei. J Comp Neurol. 1996;370(2):247-261..
9. Oyabu A, Narita M, Tashiro Y. The effects of prenatal exposure to valproic acid on the initial development of serotonergic neurons. Int J Dev Neurosci. 2013;31(3):202-208.
10. Mychasiuk R, Richards S, Nakahashi A, Kolb B, Gibb R. Effects of Rat Prenatal Exposure to Valproic Acid on Behaviour and Neuro-Anatomy. Dev Neurosci-basel. 2012;34(2-3):268-276.
11. Fueta Y, Sekino Y, Yoshida S, Kanda Y, Ueno S. Prenatal exposure to valproic acid alters the development of excitability in the postnatal rat hippocampus. Neurotoxicology. 2018.
12. Perez-Pouchoulen M, Miquel M, Saft P, Brug B, Toledo R, Hernandez ME, Manzo J. Prenatal exposure to sodium valproate alters androgen receptor expression in the developing cerebellum in a region and age specific manner in male and female rats. Int J Dev Neurosci. 2016;53:46-52.
13. Reynolds S, Millette A, Devine DP. Sensory and Motor Characterization in the Postnatal Valproate Rat Model of Autism. Dev Neurosci-basel. 2012;34(2-3):258-267.
14. Manzo J, Miquel M, Toledo R, Mayor-Mar JA, Garcia LI, Aranda-Abreu GE, Caba M, Hernandez ME. Fos expression at the cerebellum following non-contact arousal and mating behavior in male rats. Physiol Behav. 2008;93(1-2):357-363.
15. McGinnis M, Dreifuss R. Evidence for a role of testosterone-androgen receptor interactions in mediating masculine sexual behavior in male rats. Endocrinology. 1989;124(2):618-626.
16. Sachs BD, Barfield RJ. Temporal patterning of sexual behavior in the male rat. J Comp Physiol Psych. 1970;73(3):359.
17. Lucio RA, Manzo J, Martínez-Gómez M, Sachs BD, Pacheco P. Participation of pelvic nerve branches in male rat copulatory behavior. Physiology Behav. 1994;55(2):241-246.
18. Hernandez M, Soto-Cid A, Rojas F, Pascual LI, Aranda-Abreu GE, Toledo R, Garcia LI, Quintanar-Stephano A, Manzo J. Prostate response to prolactin in sexually active male rats. Reprod Biol Endocrin. 2006;4(1):1-12.
19. DeFilippis M, Wagner K. Treatment of Autism Spectrum Disorder in Children and Adolescents. Psychopharmacol Bull. 2016;46(2):18-41.
20. Anderson GM, Scahill L, McCracken JT, McDougle CJ, Aman MG, Tierney E, Arnold LE, Martin A, Katsovich L, Posey DJ, Shah B, Vitiello B. Effects of Short- and Long-Term Risperidone Treatment on Prolactin Levels in Children with Autism. Biol Psychiat. 2007;61(4):545-550.
21. McGill TE, Coughlin RC. Ejaculatory Reflex and Luteal Activity Induction In Musculus. J Reprod Fertil 1970 21: 215–220.
22. Crane LH, Martin L. Postcopulatory myometrial activity in the rat as seen by video-laparoscopy. Reprod Fertil Dev 1991 3: 685-698.
23. Yin Z, Sada AA, Reslan OM, Narula N, Khalil RA. Increased MMPs expression and decreased contraction in the rat myometrium during pregnancy and in response to prolonged stretch and sex hormones. Am J Physiol Endocrinol Metab 2012 303: 55–70.
24. Langendijk P, Bouwman E, Soede N, Taverne M, Kemp B. Myometrial activity around estrus in sows: spontaneous activity and effects of estrogens, cloprostenol, seminal plasma and clenbuterol. Theriogen 2002 57: 1563–1577
25. Waynforth HB, Flecknell PA. Experimental and Surgical Technique in the Rat Neuroscience. Academic Press, eds. London. 1999 pp 113-114.
26. Maeda KI, Ohkura S, Tsukamura H. Physiology of reproduction. En: G. J. Krinke, ed.The Laboratory Rat, Academic Press, New York. 2000 145–176.
27. Westwood FR. The Female Rat Reproductive Cycle: A Practical Histological Guide to Staging. Toxicol Pathol 2008 36: 375-384.
28. Goldman JM, Murr AS, Cooper RL. The rodent estrous cycle: characterization of vaginal cytology and its utility in toxicological studies. Birth Defects Res B Dev Reprod Toxicol 2007 80: 84–97
29. Woolley C, McEwen BS. Estradiol mediates fluctuation in hippocampal synapse density during the estrous cycle in the adult rat. J Neurosci 1992 12: 2549–2554.
30. Blume SR, Freedberg M, Vantrease JE, Chan R, Padival M, Record MJ, De Joseph MR, Urban JH, Rosenkranz JA. Sex-and estrus-dependent differences in rat basolateral amygdala. J Neurosci 2017 37: 10567-10586
31. Södersten P. Estradiol-Progesterone Interactions in the Reproductive Behavior of Female Rats. In: Ganten D., Pfaff D. (eds) Actions of Progesterone on the Brain. Current Topics in Neuroendocrinology, vol 5. Springer, Berlin, Heidelberg 1985 pp 141-174
32. Hoar W, Hickman CP. Ovariectomy and the estrous cycle of the rat. In: W. Hoar & C. P. Hickman (eds.), General and comparative physiology, 2. ed. Prentice-Hall, New Jersey. 1975 pp 260-265
33. Butcher RL, Collins WE, Fugo NW. Plasma Concentration of LH, FSH, Prolactin, Progesterone and Estradiol-17β Throughout the 4-Day Estrous Cycle of the Rat. Endocrinol 1974 94: 1704–1708.
34. Wheaton LG, Pijanowski GJ, Weston PG, Burke TJ. Uterine motility during the estrous cycle: studies in healthy bitches. Am J Vet Res 1988 49: 82-6.
35. Sheldrick EL, Flint AP. Circulating concentrations of oxytocin during the estrous cycle and early pregnancy in sheep. Prostangland 1981 22: 631-636
36. Beach FA. Sexual attractivity, proceptivity,and receptivity in female mammals. Horm Behav 1976 70: 105-138
37. Pfaff DW. Nature of sex hormone effects on rat sex behavior: specificity of effects and individual patterns of response. J Comp Physiol Psychol 1970 73: 349-358
38. Gibb W, Lye S, Challis,. Knobil, Neills. Physiology of Reproduction. In: Jimmy D. Neill (eds.), 2006 pp 2925-2974.
39. Challis JR, Lye SJ. Parturition. In: Knobil E, Neil JD, editors. The Physiology of Reproduction. New York: Raven Press 1994 pp 985–1031.
40. Shafik A. Study of the uterine response to vaginal distension: the vagino–uterine reflex. Gynecol Obstet Invest. 1997 44:265-2.
| Recibido: 04 de septiembre de 2019 | Aceptado: 17 de octubre de 2019 |
Corresponding Author at: Dr. Pedro Paredes-Ramos. Facultad de Medicina Veterinaria y Zootecnia, Universidad Veracruzana. Miguel A. de Quevedo esquina Yañez S/N, C. P 91710. Veracruz, Veracruz. México. Phone +52 (229) 9342075, ext. 24132, Fax. +52 (229) 9344053. E-mail: pparedes@uv.mx
Este es un artículo de libre acceso distribuido bajo los términos de la licencia de Creative Commons, (http://creativecommons.org/licenses/by-nc/3.0), que permite el uso no comercial, distribución y reproducción en algún medio, siempre que la obra original sea debidamente citada.