Forrajeo y polinización del arbusto Malpighia glabra (Malpighiaceae) y la orquídea Oncidium cosymbephorum (Orchidaceae) por abejas Centris (Apidae) en la isla Agaltepec, Veracruz, México
- Inicio
- Comité Editorial
- Lineamientos
- Carta de Cesión de Derechos
- Información Legal
- Acerca de la Revista
- Bases de Datos
- Contacto
- ISSN 2007-3054
- Centro de Investigaciones Cerebrales
Universidad Veracruzana
Carmona-Díaz Gustavo*1,2, García-Franco J G3, Hernández Carmona Saul1, Retureta Aponte Alejandro2, Quintana Morales Paulo C1, López Galindo Adolfo1, García Orduña Francisco1
1Instituto de Neuroetología. Universidad Veracruzana. 2Facultad de Ingeniería en Sistemas de Producción Agropecuaria. Campus Acayucan. Universidad Veracruzana. 3Instituto de Ecología, A.C.
Resumen/Abstract
Introducción
Material y métodos
Resultados
Discusión
Agradecimientos
Referencias
Correspondencia
Las abejas Neotropicales del género Centris (Apidae) están especializadas en el forrajeo de aceites florales de las especies de la familia Malpighiaceae y en la polinización de algunas especies de orquídeas del género Oncidium. La actividad diurna de las abejas Centris en las regiones tropicales de Veracruz y su grado de especialización al forrajear y polinizar solo algunos tipos particulares de flores permiten tener un excelente modelo de estudio de la ecología conductual de este singular grupo de insectos. En la región de Los Tuxtlas, Veracruz, México, se observó la interacción mutualística entre la abeja Centris lunulata, el arbusto Malpighia glabra y la orquídea Oncidium cosymbephorum. En la Isla Agaltepec, Lago de Catemaco, Municipio de Catemaco, Veracruz, se realizaron observaciones directas e indirectas de la actividad diurna de las abejas al visitar las flores de M. glabra y O. cosymbephorum. Se registró la hora de visita, el número de flores visitadas y la frecuencia, y la duración de las visitas. Se registró el sistema de reproducción de las dos plantas. Se exploró una correlación entre el horario de actividad de las abejas y la hora del día. Centris lunulata presentó alta frecuencia y duración de visitas y una mayor actividad de forrajeo y polinización en ambas especies de plantas. Se discute sobre la interacción planta-animal mediante la ecología conductual de estas abejas.
Palabras clave: Apidae, Malpighiaceae, Orchidaceae, Centris, Polinización, Forrajeo.
The Neotropical bees of the genus Centris (Apidae) are specialized in the foraging of floral oils of the species of the family Malpighiaceae and in the pollination of some species of orchids of the genus Oncidium. The diurnal activity of these bees in the tropical regions of Veracruz and their degree of specialization in foraging and pollinating only some particular types of flowers allow having an excellent model of study of the behavioral ecology of this singular group of Insects. In the region of Los Tuxtlas, Veracruz, Mexico, we observed the mutualistic interaction between the bees Centris lunulata, the shrub Malpighia glabra and the orchid Oncidium cosymbephorum. The work was done on the island Agaltepec, Lake Catemaco, municipality of Catemaco, Veracruz. We made direct and indirect observations of the diurnal activity of bees by visiting M. glabra and O. cosymbephorum flowers. Visiting time, number of visited flowers and frequency and duration of visits were recorded. In both plant species the reproduction system was recorded. A correlation between the hours of activity of bees and the time of day was explored. It was found that the bee C. lunulata on average visits greater number of flowers on both plants and has more frequency and duration of visits and greater foraging and pollination activity in both plant species. The plant-animal interaction is discussed through behavioral ecology.
Keywords: Apidae, Malpighiaceae, Orchidaceae, Centris, Pollination, Foraging.
El género Centris tiene 232 especies registradas alrededor del mundo clasificadas en tres grandes linajes (Centris, Melacentris, Trachina) lo que representa una gran diversidad de estas abejas.1 Se han reportado algunas especies de abejas de este género como visitantes florales de muchas Angiospermas y otras como sus potenciales polinizadores durante la actividad de forrajeo al recolectar los aceites o el polen.2,3 La polinización en las orquídeas ocurre cuando la abeja toca con su cabeza la capucha que contiene los polinios, adhiriéndose estos en la parte frontal de su cabeza para ser transportados hacia otras flores.4,5 En las Malpighiaceae, la polinización se presenta por el acarreo del polen en la parte ventral de una flor a otra.2-4 No obstante, la polinización por abejas en las orquídeas tiene un alto grado de especialización con sus visitantes florales y polinizadores, pudiendo llegar a depender de una o muy pocas especies de abejas para la polinización de las flores.6
Lo mismo ocurre en el caso de la polinización de las Malpighiaceae que tiene una especialización muy marcada por abejas del género Centris.7 Los escasos estudios de interacción abeja-planta en familias como Orchidaceae y Malpighiaceae, hacen esencial la identificación de polinizadores potenciales para conocer la ecología del comportamiento durante el proceso de polinización de estos dos grupos de plantas. Existe estrecha relación entre abejas del Género Centris (Apidae) y la polinización de orquídeas del género Oncidium.7 Pero pocas especies de abejas Centris han sido documentadas como polinizadoras de estas orquídeas.8 En primer lugar, porque es difícil ubicar poblaciones de las orquídeas de este género9 y en segundo lugar, porque es aún menos frecuente encontrar sitios donde habiten tanto orquídeas del género Oncidium10 como individuos de la familia Malpighiaceae.11 Las orquídeas Oncidium son epífitas y en pocas ocasiones utilizan como hospederos a árboles, arbustos o lianas de Malpighiaceae y lo más común es que se encuentren en hospederos de otras familias de plantas.12 En la región de Los Tuxtlas, Veracruz, se observó a abejas Centris13 teniendo gran actividad de visitas sobre un el arbusto Malpighia glabra, en las cuales, removían y depositaban los polinios de la orquídea Oncidium cosymbephorum que es endémica de Veracruz.14 La forma en que las abejas Centris efectúan la remoción de polinios es como se ha reportado para algunas especies de abejas del género sobre otras especies de plantas. Además de que se caracteriza por ser recolectoras de aceites florales, actividad durante la cual debe haber un adecuado ajuste entre el polinizador y la flor para establecer contacto con las partes florales reproductivas y pueda efectuarse la remoción y deposición de los polinios.12 Es importante destacar que los individuos de la orquídea O. cosymbephorum crecían de forma epífita sobre los arbustos de M. glabra, por lo cual, se pudo registrar el comportamiento del polinizador en ambas especies de plantas. Por lo anterior, el presente estudio tuvo como objetivo registrar a los polinizadores de O. cosymbephorum y cuantificar la remoción y deposición de polinios, así como también a los de M. glabra por la remoción del polen, determinando el sistema de polinización entre las abejas Centris y las flores de O. cosymbephorum y M. glabra. Las preguntas de estudio fueron las siguientes: 1) ¿Cómo es el sistema de polinización de ambas plantas? ¿Las abejas del género Centris son polinizadoras de ambas especies de plantas? ¿Cómo es la actividad de forrajeo y polinización de las abejas Centris?
El trabajo se efectuó en la Isla Agaltepec (18° 24’ - 18° 25’ de latitud Norte y 95° 05’ - 95° 06’ de longitud Oeste), Municipio de Catemaco, Veracruz, México (Figura 1). El área tiene 8.3 ha y se ubica a 432 m s. n. m. El clima es cálido-húmedo con temperatura máxima de 36°C y mínima de 13°C. La precipitación promedio es de 1980 mm. Además, la vegetación se integra por selva mediana subcaducifolia, riparia y secundaria.14 Figura 1. Isla Agaltepec, Lago de Catemaco, Municipio de Catemaco, Veracruz, México. 2.1. Autopolinización Para evaluar la necesidad de un vector (polinizador), se seleccionaron al azar 20 individuos de M. glabra y 40 de O. cosymbephorum. A cada individuo de M. glabra se le embolsaron tres ramas con malla mosquitero que contenían 113 botones florales. El control de M. glabra consistió en dejar sin bolsa a tres ramas del mismo individuo con similar número de botones. En 20 individuos de O. cosymbephorum, se embolsó con malla mosquitero una inflorescencia, conteniendo 12 flores. El control consistió en dejar sin embolsar a 20 plantas seleccionadas al azar. Los registros de producción de frutos se hicieron semanalmente y las bolsas se retiraron cuando las flores de las plantas terminaron su tiempo de vida. Se contabilizó el número de frutos producidos en los grupos controles y experimentales y se realizó una comparación. 2.2. Polinizadores El registro de los visitantes florales y potenciales polinizadores se realizó para ambas especies de plantas y las observaciones se realizaron durante la época de floración que ocurre de septiembre a diciembre. Se seleccionaron al azar los individuos y fueron observados por tres días continuos. Además, a cada visitante se le registró el número de flores visitadas, la frecuencia y la duración de la visita, y se anotó la temperatura ambiente con un termo higrómetro marca Sunto. Se exploró una correlación entre el horario de actividad de los visitantes florales y la hora del día. Las abejas polinizadoras se distinguieron de los visitantes a partir del registro de la actividad desarrollada durante la visita. Los polinizadores de O. cosymbephorum se designaron por la remoción y transporte de los polinios y/o por la deposición de los polinios en el estigma y en el caso de la planta de M. glabra fue distinguiendo la remoción, transporte y/o colocación del polen. Además, se realizaron observaciones indirectas de la remoción de polinios de O. cosymbephorum registrando las flores al final del día que quedaron sin polinios. Finalmente, se capturaron algunos especímenes utilizando una red entomológica los cuales fueron enviados para su determinación al Departamento de Química y Biología de la Universidad de Las Américas en Puebla, México y al Departamento de Entomología de la Universidad de Kansas, Estados Unidos, con los doctores Carlos Vergara Briceño y Ricardo Ayala, respectivamente.
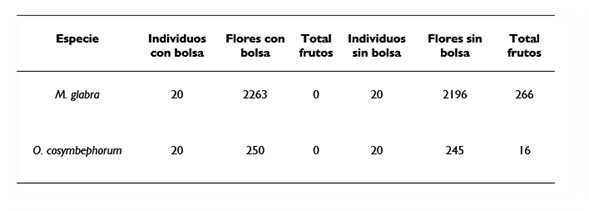
3.1. Autopolinización Las inflorescencias de ambas especies de plantas que fueron excluidas de los polinizadores no produjeron frutos a diferencia de las que estuvieron expuestas que sí desarrollaron frutos. Los individuos de M. glabra produjeron 266 frutos en total, mientras que las plantas de O. cosymbephorum desarrollaron 16 frutos (Cuadro 1). Cuadro 1. Frutos producidos de M. glabra y O. cosymbephorum en el experimento de autopolinización 3.2. Polinizadores Se registraron tres abejas de la familia Anthophoridae visitando las flores de O. cosymbephorum y M. glabra, de las cuales, Centris lunulata (Figura 2) se observó con mayor frecuencia de visita que Centris sp.1 y Centris sp.2 en flores de M. glabra (Cuadro 2). Las tres especies de abejas son recolectoras de los aceites florales producidos en los elaióforos de M. glabra. Figura 2. Abeja Centris lunulata polinizador y recolectora de aceites de las flores del arbusto Malpighia glabra (izquierda) y polinizador de la orquídea endémica de Veracruz Oncidium cosymbephorum (derecha).
Cuadro 2. Actividad de las tres abejas del género Centris en las flores de M. glabra.
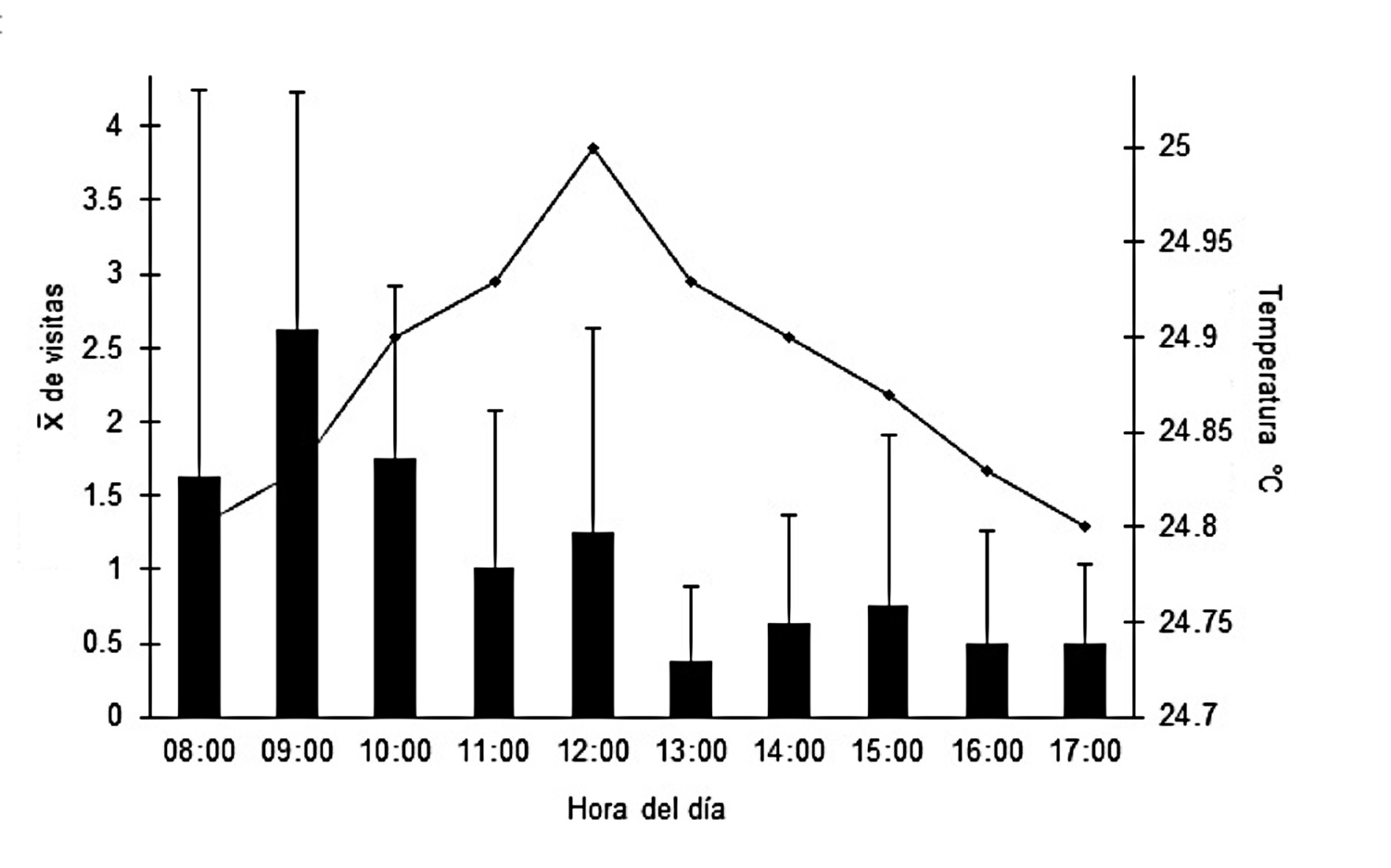
Centris lunulatatiene aproximadamente 1.5 cm de longitud y su cabeza es de color negro y el abdomen de tono anaranjado y la parte dorsal tiene una banda angosta de color amarillo, muy visible cuando se encuentran en vuelo. Al parecer son abejas solitarias que nidifican en el suelo donde hacen cavidades de aproximadamente 20 cm de profundidad. Sin embargo, poco se conoce sobre su biología. Por otro lado, Centris sp.1 tiene aproximadamente 2 cm de longitud y su cabeza es de color negro y el abdomen de tono anaranjado y no tiene banda amarilla. Probablemente esta especie es colonial, ya que frecuentemente se le observó visitando las flores de M. glabra en grupos de 3-7 individuos. También es probable que sea territorial puesto que fue más agresiva que las otras dos. Centris sp.2 es otra abeja que se observó visitando ambas especies de plantas. Esta especie fue vista en pocas ocasiones. Además, durante las visitas que efectuaron a O. cosymbephorum se registró la remoción de polinios, aunque se observó escasa actividad observada de esta abeja sobre las flores de M. glabra. Es importante indicar que no se pudo recolectar un ejemplar de esta abeja por lo cual no se determinó su especie. El patrón de actividad de C. lunulata varió en el espacio y tiempo. Las observaciones directas sobre las flores de M. glabra, indicaron que C. lunulata tuvo mayor frecuencia y duración de visitas en M. glabra que en O. cosymbephorum. La actividad de C. lunulata de acuerdo a la hora del día fue con mayor número de visitas a las flores de M. glabra entre las 08:00 y las 12:00 h, decreciendo esta actividad de las 13:00 a las 17:00 h (r = - 0.826, P <0.003) (Figura 3). Las visitas de C. lunulata a las flores de M. glabra fueron muy rápidas (menos de 1 segundo por flor), durante las cuales se observó que recolectaba el aceite de los elaióforos. Además, antes de posarse sobre una flor, C. lunulata primero la visualizaba unos cuantos segundos, para después posarse colocando la cabeza enfrente del pétalo más grande y sus extremidades entre los demás pétalos justo encima de cada par de elaióforos. De esta forma apretaba las glándulas para extraerles el aceite y almacenarlo en sus tibias, en esta postura el polen se adhería al centro del abdomen.
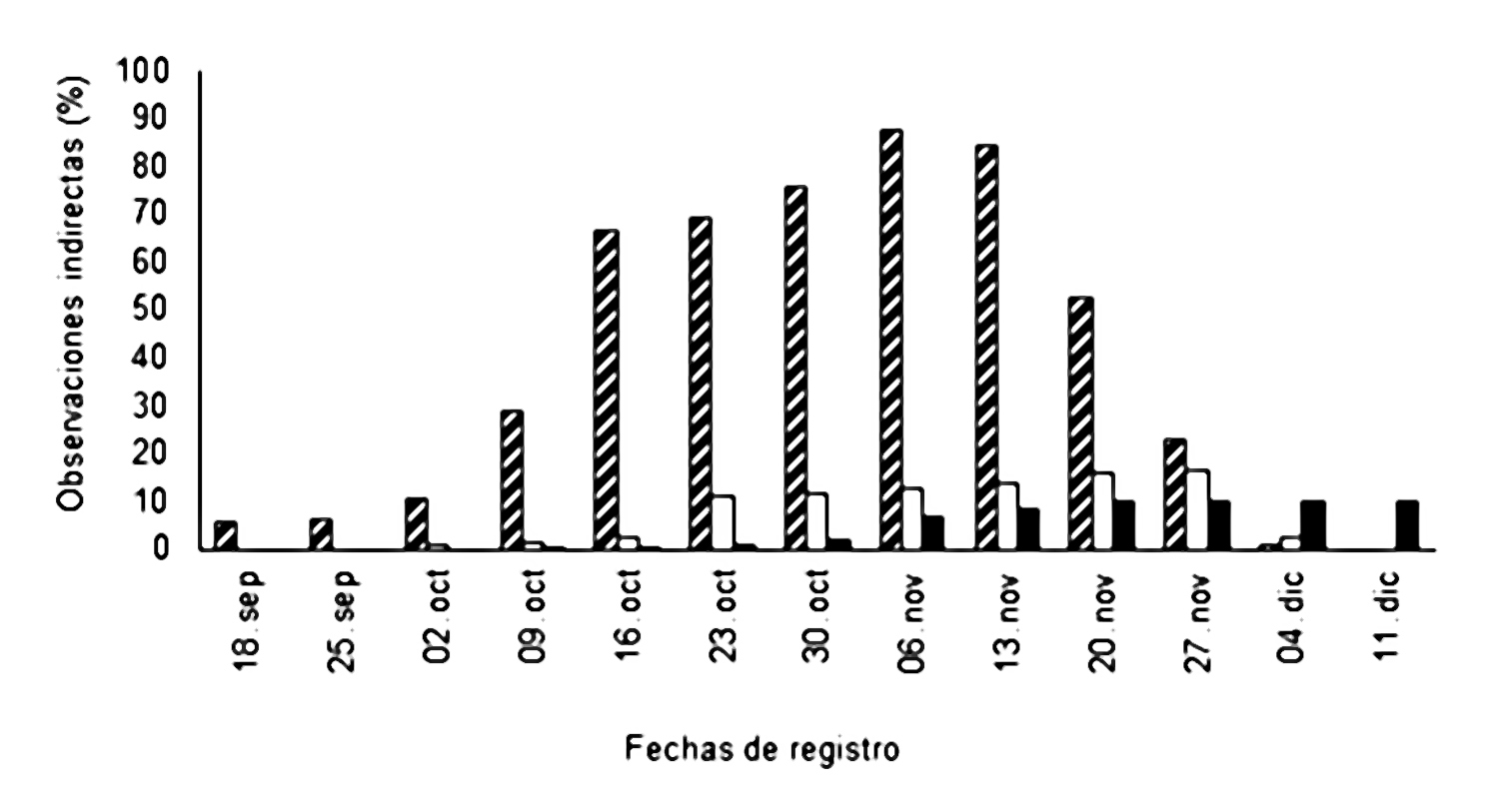
En cuanto a la actividad de C. lunulata sobre las flores de O. cosymbephorum, se observó a C. lunulata removiendo y depositando polinios en las flores de otras plantas de O. cosymbephorum que resultaron en la producción de frutos. Las observaciones indirectas de O. cosymbephorum mostraron 981 flores sin remoción de polinios, 141 con remoción y 115 frutos producidos cuyo porcentaje se muestra en la Figura 4. Es pertinente indicar que para la deposición de los polinios es necesaria una visita doble por parte de las abejas: 1) al retirarlos de una flor y 2) al colocarlos en la flor de otra planta.
Figura 3. Actividad de Centris lunulata al efectuar el forrajeo de las flores de Malpighia glabra a lo largo del día. La línea indica la temperatura que se registró durante los días de observación.
Figura 4. Observaciones indirectas de la actividad de Centris lunulata sobre las flores de Oncidium cosymbephorum para remover y depositar los polinios. (barras transversales = flores con polinios; barras blancas = flores con polinios removidos; barras negras = frutos).
La actividad de forrajeo de las abejas C. lunulata durante la polinización de M. glabra y O. cosymbephorum es evidencia de que ambas plantas necesitan de uno o varios polinizadores para la fecundación de sus flores, puesto que las flores excluidas de las dos especies de plantas no produjeron frutos. La transferencia del polen en M. glabra y de los polinios en O. cosymbephorum debe ser efectuada por C. lunulata, por lo cual, las plantas estudiadas no son autógamas y requieren de polinizadores. La estrecha relación mutualística entre Malpighiaceae y Apidae-Centridinii mencionada en la literatura hacía suponer que abejas de los géneros Centris, Epicharis o Hemisiella1,2,5 fueran los visitantes florales y potenciales polinizadores de Malpighia glabra. Sin embargo, en algunas especies arbustivas del género Malpighia se conoce que abejas recolectoras de aceites del género Centris actúan como únicos visitantes florales, infiriendo que por tal razón, deberían ser también los potenciales polinizadores en la especie estudiada.5 Los registros sobre estas especies de abejas corresponden a observaciones aisladas y anecdóticas de las visitas de estas abejas a las flores de Malpighia, sin efectuar un estudio particular sobre la polinización, la autopolinización de las plantas y la actividad de forrajeo de las abejas. Particularmente, el comportamiento observado de Centris lunulata muestra la importancia de considerar la actividad de la alimentación junto con la polinización. En algunos estudios se ha mostrado que las abejas Centris pueden modificar su comportamiento de alimentación, y, por lo tanto, de polinización dependiendo de las condiciones climáticas y fenológicas.5,13,14 Sin embargo, la escasa presencia de individuos de C. lunulata, se refleja en la poca producción de frutos de M. glabra y nula presencia de frutos de O. cosymbephorum.14 La actividad diurna de C. lunulata con mayor actividad antes del mediodía (ver Figura 3) puede tener algunas posibles explicaciones. La primera tiene que ver con la temperatura que influye directamente con la actividad de cualquier abeja.4 Muchas especies de abejas tienen óptimos de temperatura donde desarrollan sus múltiples actividades como la alimentación y la construcción de nidos.6 En el caso de C. lunulata probablemente el aumento de la temperatura y de la sensación térmica que puede subir varios grados más, implique una disminución de actividad a lo largo del día por el sobrecalentamiento corporal de los individuos. En varias ocasiones se observaron individuos de esta abeja en días nublados y con lluvia con una mayor frecuencia de visita que en días soleados, aunque esto también puede reflejar la necesidad de materiales para alimentación o construcción y el cuidado parental.15 Otra posible explicación es que por el aumento de la temperatura, los aceites esenciales florales de M. glabra se dejen de producir en los elaióforos del arbusto, ya bien sea por un mecanismo fisiológico de la planta o por evaporación con lo cual las abejas C. lunulata dejan de visitar por la falta de recompensa floral.16 Para la orquídea O. cosymbephorum no se tenían registros de las abejas que removían y depositaban polinios en los estigmas para la fecundación y producción de frutos. Siendo O. cosymbephorum una especie sin recompensa floral como aroma, aceites esenciales volátiles o néctar14 se proponía como autógama por lo que la presencia de polinizadores no era necesaria o bien que la frecuencia de visita y actividad de forrajeo fuera muy escasa. Las visitas frecuentes, la remoción y deposición de polinios evidenciaron que C. lunulata es el polinizador potencial de O. cosymbephorum. Aunque no se descarta que, en años posteriores, las otras dos especies de las abejas Centris sp.1 y Centris sp.2, puedan presentar mayor número poblacional en comparación con Centris lunulata y funcionar como polinizadores de M. glabra y O. cosymbephorum. Desde nuestro entendimiento no hay registrado una interacción donde tres abejas del género Centris participen activamente en el forrajeo y la polinización de dos especies de plantas de diferentes familias, pero con morfología y color florales muy similares. El hecho de que M. glabra con recompensa floral y O. cosymbephorum sin recompensa floral compartan el mismo polinizador a pesar de la diferencia en el tamaño de las flores, muestra que otros factores deben estar involucrados en la atracción de C. lunulata. La morfología floral entre ambas plantas no es idéntica, pero si existen similitudes entre sus partes florales, por lo que algunos rasgos en las flores (estructuras y/o colores), pueden ser los detonadores de la interacción tripartita. La similitud entre las flores de M. glabra y O. cosymbephorum parece tener varios componentes. Por un lado, las estructuras florales fueron similares (pétalos y sépalos), aunque también están relacionados morfológicamente los elaióforos y el polen de M. glabra con las verrugosidades del labelo y los polinios de O. cosymbephorum. El color es un componente importante en la atracción de C. lunulata y en M. glabra y O. cosymbephorum es probable que el color magenta sea el principal responsable en la atracción de C. lunulata. Hasta el momento no hay claridad sobre las características florales involucradas directamente en la atracción de C. lunulata, por lo que serán importantes las manipulaciones experimentales en la simetría y morfología floral y pigmentación de las estructuras florales en ambas especies.
La forma biológica epífita de O. cosymbephorum permitió observar que la distancia entre esta y M. glabra es determinante para que Centris lunulata visite las flores de la orquídea. Las inflorescencias de O. cosymbephorum que estaban cerca de las ramas de M. glabra presentaron mayor cantidad de frutos en comparación con aquellas donde la distancia con M. glabra fue más grande. Se observó que, sin la presencia de M. glabra, el éxito reproductivo de la orquídea parece disminuir, aunque esto debe probarse de manera experimental, ya que la similitud estructural floral puede hacer posible que favorezca la polinización. En este caso existe la posibilidad de que la similaridad14 de O. cosymbephorum sea adaptativa17 y que la presencia de las flores del arbusto M. glabra este directamente implicada en el éxito reproductivo de O. cosymbephorum. Esto hace suponer que el sistema de polinización de M. glabra es por la recompensa floral (polen y aceites) que obtiene C. lunulata pero que la polinización de O. cosymbephorum con flores sin recompensa floral implique otro tipo de sistema puesto que aún sin la recompensa parece implicar la atracción de C. lunulata. Dos hipótesis deberán ser probadas, mimetismo floral batesiano y efecto magnético de las especies. Pero también resulta interesante mostrar que el éxito reproductivo de O. cosymbephorum junto a M. glabra solo refleje la alta densidad de polinizadores visitando más a M. glabra por las recompensas, pero esto tendría que probarse experimentalmente en poblaciones aisladas donde la orquídea se encuentre sin la presencia de M. glabra.
Los autores agradecen a Ramón Santos Villegas su apoyo en campo y a los tres revisores anónimos por sus atinadas observaciones. Los autores declaran no tener conflictos de intereses. Gustavo Carmona Díaz. Instituto de Neuroetología. Universidad Veracruzana. Avenida Dr. Luis Castelazo, Industrial de Las Ánimas, 91190, Xalapa, Veracruz, México. Tel: +52 (228). 8421700. Ext. 11708. Correo electrónico: floralmimicry@yahoo.com.mx Este es un artículo de libre acceso distribuido bajo los términos de la licencia de Creative Commons, (http://creativecommons.org/licenses/by-nc/3.0), que permite el uso no comercial, distribución y reproducción en algún medio, siempre que la obra original sea debidamente citada. 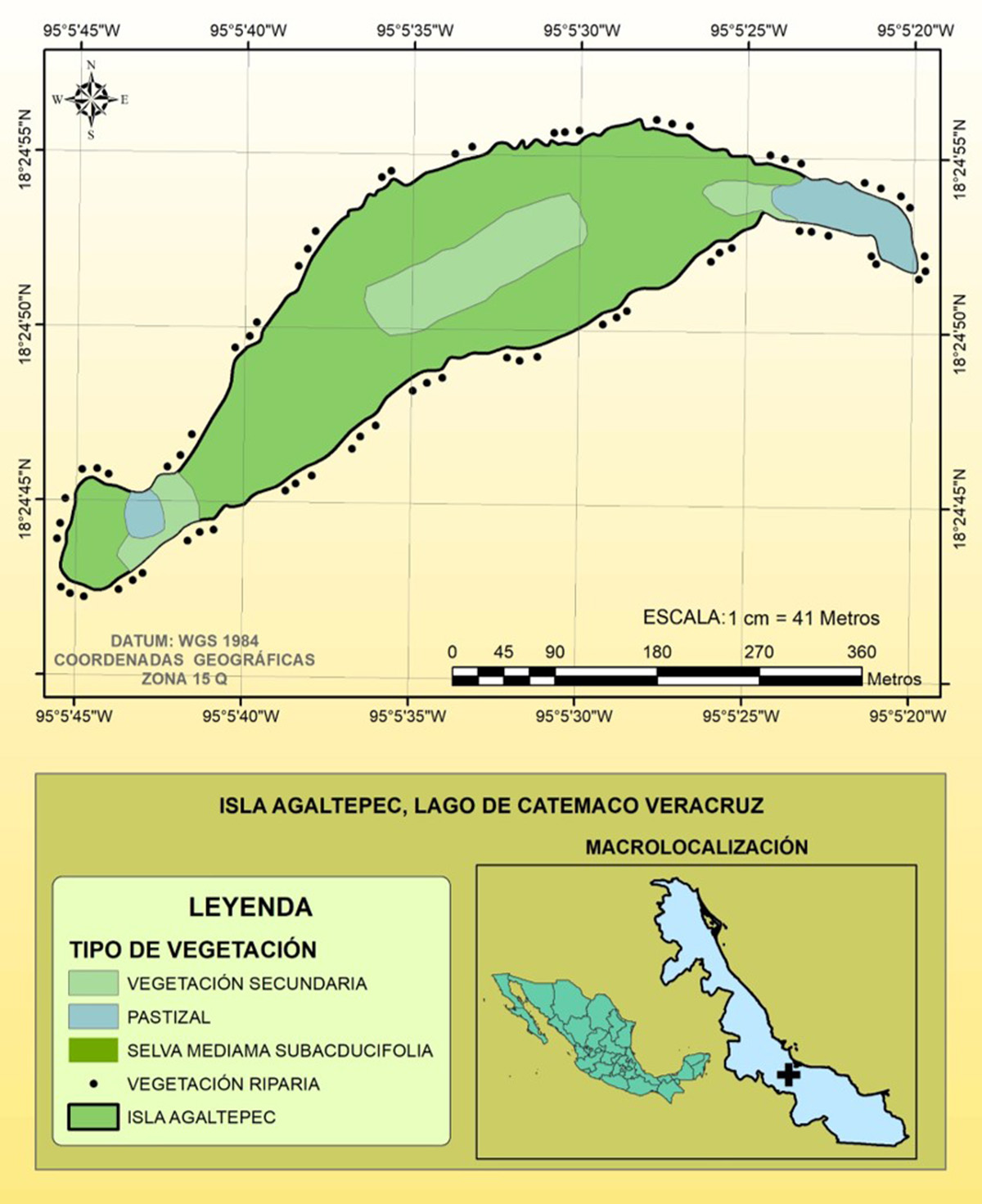


Recibido: 19 de septiembre de 2017
Aceptado: 18 de diciembre de 2017