Selective deprivation of rapid eye movement (REM) sleep for 24 h does not modify the c-Fos immunoreactivity in the Ventral Respiratory Column (VRC) of rat
- Inicio
- Comité Editorial
- Lineamientos
- Carta de Cesión de Derechos
- Información Legal
- Acerca de la Revista
- Bases de Datos
- Contacto
- ISSN 2007-3054
- Centro de Investigaciones Cerebrales
Universidad Veracruzana
Muñoz-Ortiz Elizabeth1†, Díaz-Escárcega Rocío1†, Meza-Andrade Roberto2, Muñoz-Ortiz Janeth1, García-García Fabio3, López-Meraz Leonor2, Beltran-Parrazal Luis2, Morgado-Valle Consuelo2*
1Doctorado en Investigaciones Cerebrales, Universidad Veracruzana. 2Centro de Investigaciones Cerebrales, Universidad Veracruzana. 3Instituto de Ciencias de la Salud, Universidad Veracruzana.
†These authors contributed equally.
Resumen/Abstract
Introducción
Material y métodos
Resultados
Discusión
Agradecimientos
Referencias
Correspondencia
La generación y modulación de la respiración es controlada por la columna respiratoria ventral (VRC), la cual incluye: el grupo respiratorio parafacial/núcleo retrotrapezoide (pFRG/RTN), el complejo Bötzinger (BötC) y el complejo preBötzinger (preBötC). Una correlación entre el sueño y la respiración es particularmente clara en condiciones como la apnea obstructiva del sueño y el síndrome de hipoventilación central congénita. En este trabajo, mediante la inmunoreactividad contra c-Fos como un marcador indirecto de la actividad neuronal, se estudió el efecto de la privación selectiva de sueño MOR a corto plazo sobre la actividad de las neuronas en la VRC. Ratas macho Wistar fueron divididas en dos grupos: control (n=6) y 24 h de privación selectiva de sueño MOR utilizando la técnica de florero invertido (n=11). La frecuencia respiratoria, la frecuencia cardíaca, la temperatura y la saturación de oxígeno (SpO2) se midieron antes y durante la privación de sueño MOR. El grupo de privación de sueño MOR no mostró diferencias estadísticamente significativas con respecto al grupo control. Sin embargo, la frecuencia cardíaca del grupo de privación de sueño MOR aumentó y la SpO2 disminuyó significativamente en comparación con los valores basales. Estos datos sugieren que, en ausencia de sueño MOR, la actividad de las neuronas respiratorias en la CRV está altamente regulada para asegurar la homeostasis de gases y una tasa de respiración estable y que los cambios fisiológicos encontrados probablemente están regulados por núcleos no relacionados con la generación del ritmo respiratorio.
Palabras clave: Columna respiratoria ventral, Complejo preBötzinger, Ritmo respiratorio, c-Fos; Privación de sueño, Sueño MOR.
Breathing generation and modulation is controlled by several regions in the ventral respiratory column (VRC), such as the parafacial respiratory group/retrotrapezoid nucleus (pFRG/RTN), the Bötzinger Complex (BötC) and the preBötzinger Complex (preBötC). A correlation between sleep and breathing is particularly clear in conditions such as obstructive sleep apnea and congenital central hypoventilation syndrome. Here, by using c-Fos immunoreactivity as an indirect marker of neuronal activity, we aimed to study the effect of short-term selective REM sleep deprivation on the activity of neurons along the VRC. Male Wistar rats were divided in two groups: control (n=6) and 24 h selective REM sleep deprivation using the flowerpot technique (n=11). Respiratory rate, heart rate, temperature and oxygen saturation (SpO2) were measured before and during REM sleep deprivation. When comparing the REM sleep-deprived to the control group, not statistically significant differences were found in the number of c-Fos positive neurons in the pFRG/RTN and VRC or in the respiratory rate, heart rate, temperature and SpO2. Interestingly, in the REM sleep-deprived group the heart rate increased and SpO2 decreased statistically significantly compared to baseline values. These data suggest that, in the absence of REM sleep, activity of respiratory neurons in the VRC is highly regulated to ensure a stable breathing rate and gas homeostasis, and that physiological changes may result from modulation on nuclei not involved in the generation of respiratory rate.
Key words: Ventral respiratory column, preBötzinger complex, respiratory rhythm, c-Fos, sleep deprivation, REM sleep.
Respiratory sleep disorders are common in neurological and neuromuscular disorders due to a combination of factors such as muscle weakness and/or damage to areas of the brain that control breathing. Such disorders can occur predominantly in REM sleep due to a decreased muscle tone and a decreased responsiveness of chemoreceptors.1,2
In mammals, breathing is a continuous process to maintain gas homeostasis. Respiratory rhythm is generated and modulated by neural networks located in the ventral respiratory column (VRC). VRC consists of two bilateral columns along the ventrolateral medulla containing several types of respiratory neurons, which are identified by their activation during the inspiratory or expiratory phases of the respiratory cycle. Nuclei in the VRC include, from rostral to caudal: the retrotrapezoid nucleus (RTN), which is partially overlapped with the parafacial respiratory group (pFRG), the Bötzinger complex (BötC), the preBötzinger complex (preBötC) and the ventral respiratory group (VRG), which is subdivided into rostral and caudal.3-6 The preBötC plays a critical role in generating the inspiratory phase of respiratory rhythm.7
In in vivo studies in adult rats, ablation of preBötC neurons expressing the receptor to neurokinin 1 (NK1R) results in progressive and increasingly severe disruption of respiratory rate, initially during sleep.8 One important structure involved in the modulation of respiration is the pFRG/RTN, a chemosensitive structure. The contribution of pFRG/RTN to central respiratory chemoreception may be behavioral and/or state dependent.9,10
Sleep consists of two phases: of rapid-eye movements (REM) and of slow-waves or non-REM.11 In the non-REM phase, the muscle tone diminishes but does not disappear, whereas in the REM phase the muscle tone disappears. During the REM phase, there are variations in heart and respiratory rates, therefore respiration is considered fragile.12,13 During REM sleep in both, rats14 and elderly humans,15 breathing is more irregular and there are more spontaneous central apneas.
Here, we used c-Fos immunoreactivity as an indirect marker of neuronal activity to investigate whether selective deprivation of REM sleep affects neuronal activity in the brainstem, specifically in areas related to respiration.
2.1. Animals
All experimental protocols were approved by the Institutional Animal Care and Use Committee of the Universidad Veracruzana according to the Official Mexican Standard NOM-062-ZOO-1999 (Technical Specifications for the Production, Care and Use of Laboratory Animals) and we followed the National Institutes of Health guidelines for animal care and handling. Adult male Wistar rats (308.4±26.9 grams) were individually housed in a room at 24 degrees Celsius (ºC), under 12 h light/dark cycles (lights on at 8:00 AM) with water and food provided ad libitum.
2.2. REM sleep deprivation
We used the flowerpot technique for REM sleep deprivation during 24 h, starting in the light phase. Briefly, each rat (n=11) was placed on a 6.5 cm diameter circular platform in the center of a small water tub surrounded by water up to 1 cm below the surface of the platform. This set-up allows the rat to transit from wake to non-REM sleep but prevents REM sleep. As the muscle tone decrease during REM sleep the animal fall into the water and wakening. Control rats (n=6) were placed in a similar set-up on a 15 cm diameter platform, large enough to allowing the animal to transit to REM sleep without falling into the water. Control group is aimed to exclude the non-specific effects of the flowerpot technique and is considered a better control for the REM deprivation group.16 Both groups had free access to water and food during the 24 hour period in the experimental set-up.
2.3. Vital signs measurement
Respiratory rate, heart rate, rectal temperature and oxygen saturation (expressed as percentage), were measured in both groups before and during REM sleep deprivation using a non-invasive automatic electronic monitoring system (PLUTO, Biosonic Co. LTD. Korea). Measurements were taken approximately every 6 hours. Four measurements were done before placing the rat in the flowerpot set-up and four measurements during the 24 h subsequent. Baseline values were calculated by averaging the measurements previous to the flowerpot set-up from all rats (n=17).
2.4. Tissue processing
After 24 h in the flowerpot set-up, control and REM sleep deprived rats were anesthetized with a lethal dose of sodium pentobarbital and transcardially perfused with 150 ml of 0.9% saline, followed by 4% paraformaldehyde in phosphate buffer (PB, pH 7.4; 250 ml). Brainstem was obtained and post-fixed for 24 h, cryoprotected in 25% sucrose PBS and embedded in Tissue-Tek O.C.T. compound tissue-freezing medium (Sakura Finetek USA Inc.). Coronal sections of a 40 μm thickness were obtained from -11 to -14.6 mm with respect to bregma,17 considering the approximate location of the following regions: pFRG/RTN: -11.76 mm; Bötzinger: -12.36 mm; preBötC: -12.72 mm; rVRG: -13.20 mm and cVRG: -14.28 mm. Sections were incubated in rabbit anti c-Fos (Santa Cruz Biotechnology, Santa Cruz, USA) primary antibody diluted (1:2000) in serum at 4°C overnight, placed in biotin conjugated species-specific secondary antibody (Vector Laboratories, Burlingame, CA, USA), stained using the ABC method (Vector Laboratories) and mounted on gelatin-subbed slides.
2.5. Data analysis
Digital images were obtained with an optical microscope (Olympus AX70) and a 4x lens. Bilateral counts of positive immunoreactivity to c-Fos (c-Fos+) in the regions of interest were restricted to a region of interest (ROI) consisting of a circle of 600 μm in diameter (area=0.28 mm2) as in McKay et al, (2008)18 (Figure 1), using the digital image processing software, ImageJ.19 Due to the size and arrangement of the region corresponding to the pFRG/RTN, the same circle was arranged in such manner that only half (area=0.14 mm2) enclosed the tissue. c-Fos+ cells were considered to have a brown marking on the nucleus (Figure 2). Cells with labeling only in the cytoplasm were not counted. The first plane of three sections, each intercalated every 80 μm, was counted per anatomical region of interest per animal. The counting was performed in a double-blind protocol by duplicate.
The cell count, respiratory rate, heart rate, temperature and SpO2 were expressed as mean±standard error of the mean (S.E.M.). Since our sample passed normality and equal variance tests, we performed t-student tests among control group and REM-sleep deprivation group and for each group against baseline. We used Sigma Plot software.
Using c-Fos immunostaining as an indirect measurement of cellular activity, we quantified the number of c-Fos+ cells in coronal brainstem sections from -11 mm to -14.6 mm with respect to bregma15 that include the pFRG/RTN and the VRC from REM-sleep deprived and control rats (Figures 1-3).
We did not found statistically significant changes in the total number of c Fos+ cells in the VRC per rat in the REM sleep deprivation group (83±14 c-Fos+ cells; n=11) with respect to control group (71±8 c-Fos+ cells; n=6; Figure 3). Not statistically significant differences were found when comparing the number of c-Fos+ cells in each region of interest for control group and REM sleep deprivation group. Total number of c-Fos+ cells at the level of the pFRG/RTN was 9±2.5 and 6.3±3.1 (p=0.3) in control and REM sleep deprivation group, respectively. Total number of c-Fos+ cells at the level corresponding to the preBötC was 11.5±2.4 and 11±2.0 (p=0.9) in control and REM sleep deprivation group, respectively (Figure 3).
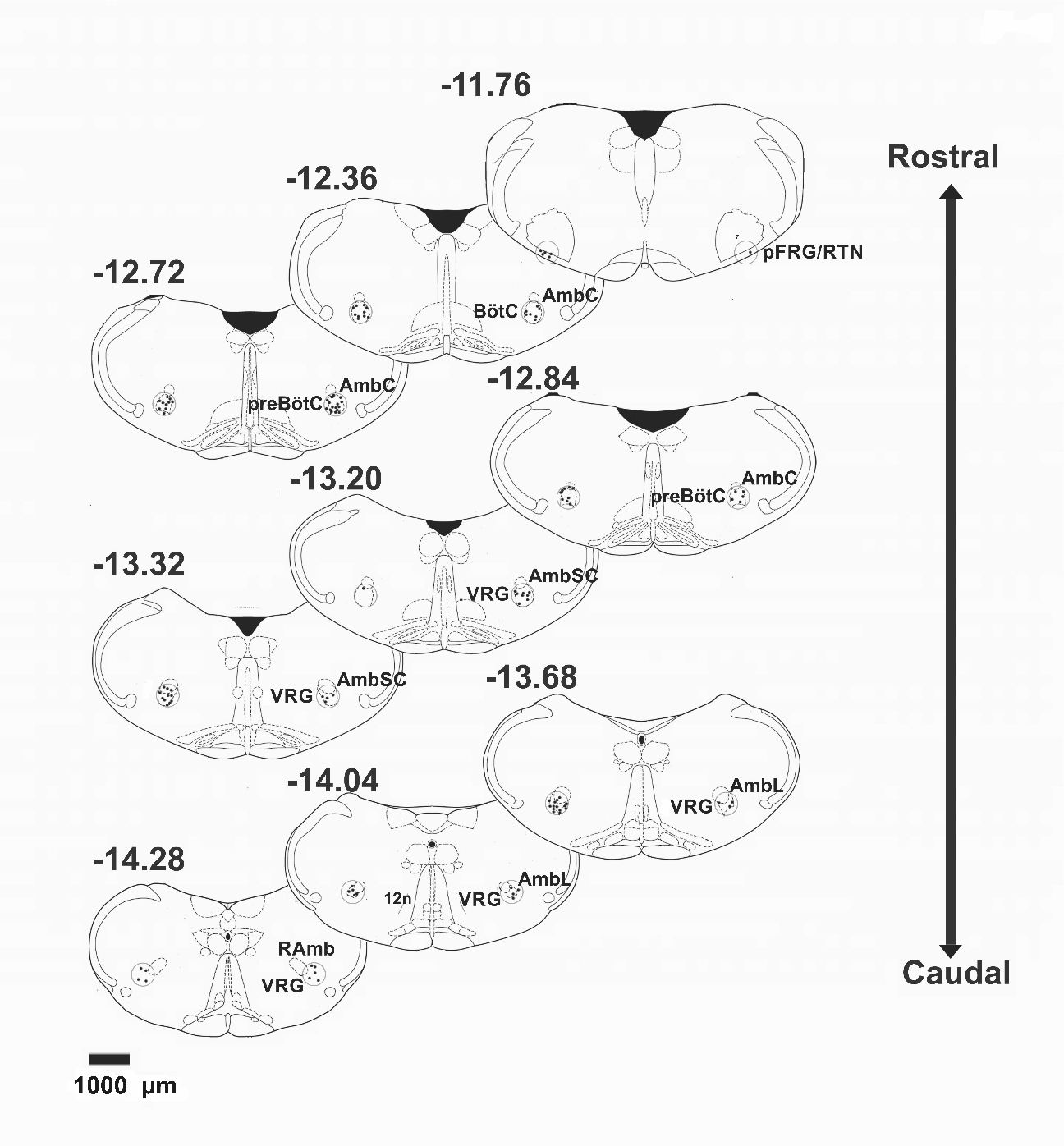
Figure 1. Schematic representation of stereotaxic levels corresponding to the parafacial respiratory group/retrotrapezoid nucleus (pFRG/RTN) and ventral respiratory column (VRC). Black circles (600 μm in diameter) represent the area where c-Fos+ cells were quantified. Numbers represent the position of the sections with respect to bregma (in mm). 7: facial nucleus. AmbC: ambiguus nucleus compact part. BötC: Bötzinger complex. PreBöC: preBötzinger complex. AmbSC: ambiguus nucleus subcompact part. VRG: ventral respiratory group. AmbL: Ambiguus nucleus loose part. 12n: hypoglossal nerve. RAmb: nucleus retroambiguus.
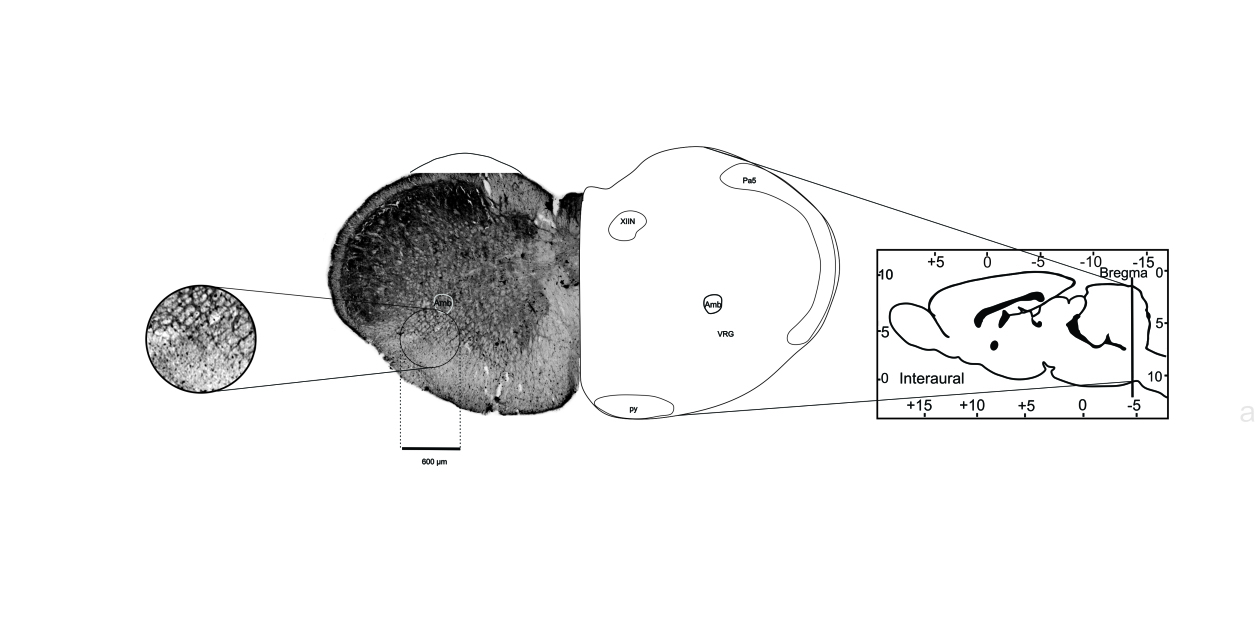
Figure 2. Immunoreactivity to c-Fos in brainstem neurons. Representative photomicrograph of a transverse slice of the brainstem of rat. The circle represents the ROI (600 µm in diameter) where c-Fos+ cells were counted. The images were obtained with a 4x objective in an optical microscope. XIIN: hypoglossal nucleus. AmbL: ambiguus nucleus loose part. VRG: ventral respiratory group. Pa5: paratrigeminal nucleus. Py: pyramidal tract.
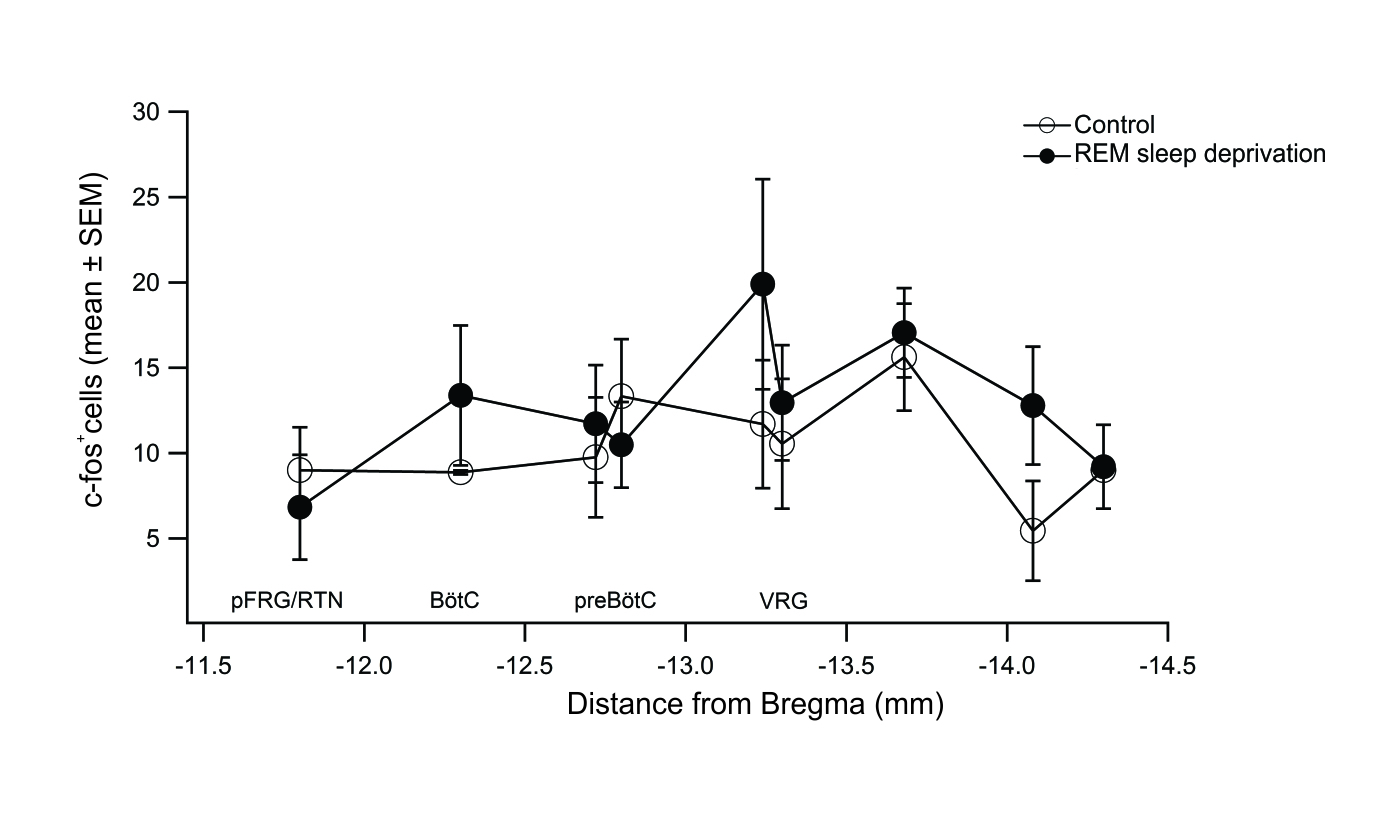
Figure 3. Rostrocaudal distribution of c-Fos+ cells in regions of the brainstem. Mean number of c-Fos+ cells per region and the approximate distance (in mm) relative to bregma of the sections counted. Control group (open circles, n=6). REM sleep deprivation group (filled circles, n=11). pFRG/RTN: parafacial respiratory group/retrotrapezoid nucleus. BötC: Bötzinger complex. PreBöC: preBötzinger complex. VRG: ventral respiratory group.
Baseline respiratory rate was 86.9±2.1 breaths per minute (bpm). During the 24 h period in the flowerpot set-up, not statistically significant changes in respiratory rate were found in the REM-sleep group with respect to control group or to baseline values. After 24 h in the flowerpot set-up, the respiratory rate in control group was 69.5±4.5 bpm and in the REM-sleep deprivation group was 79±4.9 bpm (n.s.). We found a statistically significant decrease in the respiratory rate in the control group with respect to baseline values (p<0.05, Figure 4A).
Baseline heart rate was 339.4±15.0 beats per minute (hbpm). During the 24 h in the flowerpot set-up, we did not find statistically differences in the REM-sleep deprivation group with respect to control group (Figure 4B). In both groups, although not statistically significant, we observed an increase in the heart rate with respect to baseline values after 12 h in the flowerpot set-up, which coincides with the change of phase from light to dark. After 24 h in the flowerpot set-up, the mean heart rate in control group was 358.3±31.7 hbpm and in the REM-sleep deprivation group was 411.2±20.9 hbpm (n.s.). The increase in heart rate in the REM-sleep deprivation group was statistically significant with respect to baseline values (p<0.05, Figure 4B).
Baseline temperature was 34.4±0.6 °C. We did not find statistically significant changes in the temperature in REM-sleep deprivation and control groups at any time during the 24 h period spent in the flowerpot set-up (Figure 4C). In our conditions, baseline oxygen saturation was 92.2±0.7%. We did not find statistically significant changes in SpO2 in the REM-sleep deprivation and control groups at any time during the 24 h period spent in the flowerpot set-up. However, at the end of the 24 h in the flowerpot set-up, we found a statistically significant decrease in SpO2 in the REM-sleep deprivation group (49.4±20.8%) with respect to baseline values (p<0.01, Figure 4D).
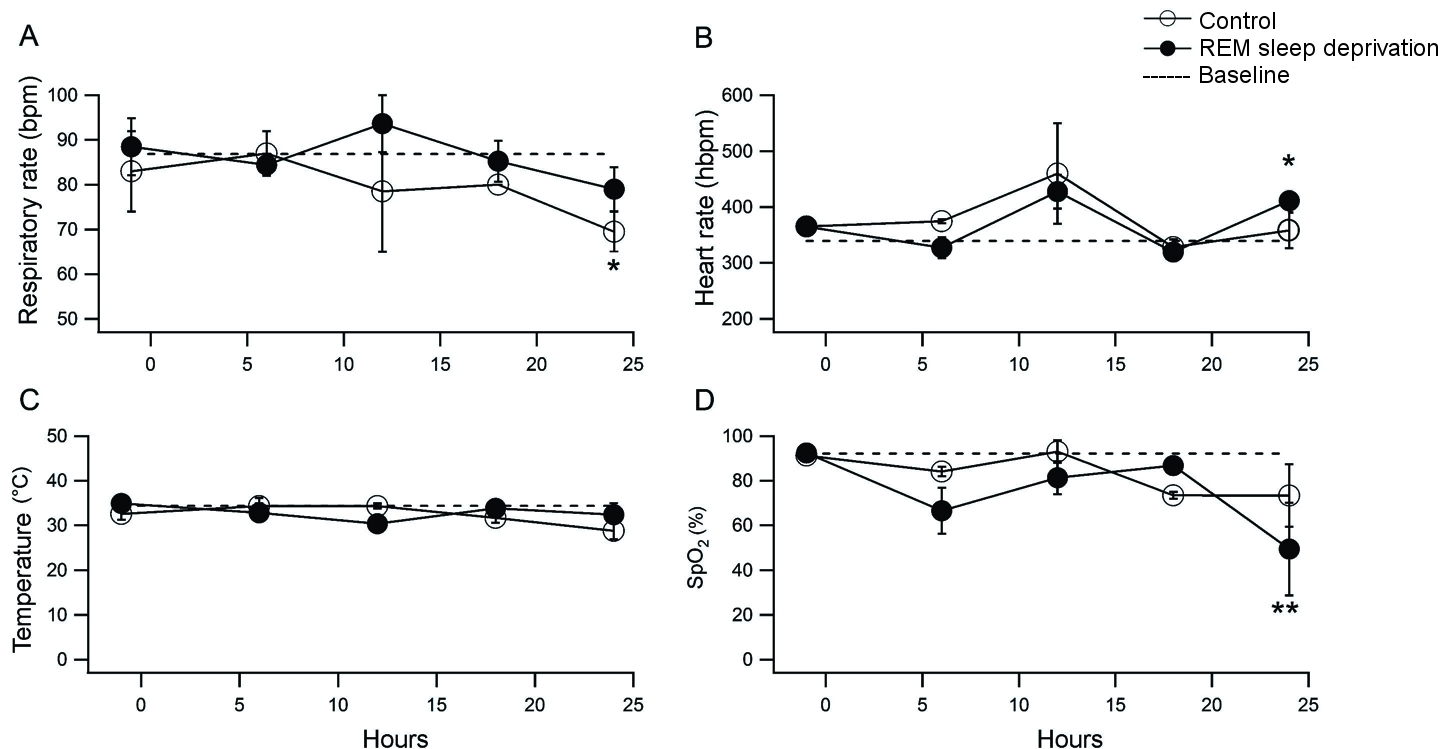
Figura 4. Effect of REM sleep deprivation on respiratory rate, heart rate, temperature and oxygen saturation (SpO2). Control group (open circles, n=6) and REM sleep deprived group (filled circles, n=11). The dotted line represents the mean of baseline measurements, prior to experimental manipulation. A. Effect of REM sleep deprivation on respiratory rate expressed in breaths per minute (bpm). * p<0.05 control vs baseline. B. Heart rate plotted in beats per minute (hbpm). C. Temperature in °C. D. Oxygen saturation reported as %. ** p<0.01 REM sleep deprivation vs baseline.
By using c-Fos immunoreactivity as a marker of cellular activity, we explored the effect of 24 h REM-sleep deprivation on brainstem regions that are involved in generation/modulation of breathing. Our data suggest that REM-sleep deprivation for 24 h does not modify the neuronal activity of VRC regions, i.e., it does not modify the activity of neurons in structures related to the generation of the inspiratory and expiratory phases of respiration.
There are few studies quantifying respiratory and cardiac rates during REM sleep deprivation in rats. Radulovacki et al., (1997)20 did not find significant changes in the heartbeat period before and after 48 h of REM sleep deprivation, but found a significant decrease in blood pressure and respiratory rate. Under our conditions, REM sleep deprivation produces changes in heart rate and SpO2 that may be due to direct modulation of motor and/or chemosensitive nuclei not present in the VRC, e.g., hypoglossal nucleus, phrenic motor nucleus, nucleus of the solitary tract and raphe nuclei. However, to testing this hypothesis requires a detailed study of the effect of REM sleep deprivation on these nuclei.
4.1. Temporal resolution of c-Fos immunoreactivity
The immunoreactivity against c-Fos as a marker of neuronal activity is a widely used tool for the study of sleep to understanding the involvement of neuronal populations during phase change, sleep deprivation and/or sleep structure.21-23 In neurons, the c-Fos mRNA reaches peak levels at 30 minutes after the stimulus and the amount of protein is maximal between 60-90 min and persists for 2-5 h.24 Expression of c-Fos after sleep deprivation has been characterized mainly in pontine and suprapontine structures. In most areas, the highest levels of c-Fos expression are seen after 3 h of sleep deprivation.22-25 There is a possibility that after 24 h of REM sleep deprivation we missed the maximum expression of c-Fos in the VRC. However, since we did not find significant changes in respiratory rate or SpO2 in the REM sleep deprivation group compared to control group, we assume that the flowerpot technique does not act as an acute stimulus in the regions of interest. An additional limitation of this study is that quantification was done after only 24 h of REM sleep deprivation. Would a prolonged REM sleep deprivation, for example for 48 h, increase c-Fos expression in VRC? Longer periods of REM sleep deprivation have a significant effect on physiology and, therefore, could lead to confusing patterns of c-Fos expression.26 More studies are needed to establish the temporal course of c-Fos expression in VRC during sleep deprivation.
Is the flowerpot technique the best model for studying REM sleep deprivation?
The flowerpot technique is the most widely used method in rodents for selective deprivation of REM sleep. This method ensures almost total deprivation of REM sleep, while the amount of non-REM sleep is reduced by approximately 10%. However, an important caveat is that this method induces stress and a unique behavioral phenotype in which there is an increase in exploratory activity, a reduction of fear and, in experiments in which the rat can self-administer voltage by pressing a bar, an increase in lateral hypothalamic self-stimulation is observed.27 In order to reduce the stress component, other methods have been developed, for example direct stimulation of the reticular formation of the mesencephalon,27 disk on water28 and treadmill.29 However, the main disadvantage of these methods is that complex instrumentation requiring feedback by electroencephalographic recording.
In an effort to quantify stress, some groups have determined the circulating levels of corticosterone before and after sleep deprivation. Sleep deprivation by gentle handling for 6 h increases 10 times the levels of corticosterone.30 REM sleep deprivation for 96 h using the multiple-platform increases 10 times the levels of corticosterone compared to control and suppresses cell proliferation in the dentate gyrus.31 In contrast, total sleep deprivation for 96 h using the treadmill method does not increase the levels of corticosterone compared to control conditions, but significantly reduces proliferation in the dorsal hippocampus.32 These data suggest that, despite the fact that sleep deprivation induces stress, the effects seen in discrete brain areas not related to the limbic system, are due to sleep deprivation and not due to stress.
4.2. Sleep and breathing
Obstructive sleep apnea is the most studied relationship between sleep and breathing. In humans, during sleep under normal conditions there is a depression in basal ventilation per minute. In the past, such depression was attributed exclusively to impaired activity of motor neurons controlling the upper airway. However, recent evidence suggests that there are central mechanisms involved in the regulation of breathing during sleep.33 Unilateral ablation of preBötC neurons expressing the NK1R alters the respiratory pattern during REM sleep in rats.18 This suggests that during REM sleep under normal conditions, the activity of preBötC neurons is depressed. In cats, the activity of respiratory neurons in the ventral medulla oblongata decreases during REM sleep.34
RTN appear to be a critical center for regulation of CO2 during breathing in a state-dependent manner.35 The RTN regulates both respiratory rate and tidal volume in states such as anesthesia, non-REM sleep, or quiet wakefulness. However, the RTN does not control the respiratory rate during REM sleep.36 Although we did not find changes in c-Fos expression in pFRG/RTN and VRC, we do not rule out the possibility that REM sleep deprivation may affect the activity of respiratory neurons in pontine structures, e.g., the pontine respiratory group (PRG). Respiratory neurons in PRG are not essential for the generation of respiratory rhythm, but it has been proposed that, along with vagal afferents, they modulate the respiratory phase change.37
4.3. Caveats
Here, we aimed to determine the impact of REM sleep deprivation on the activity of the VRC, which is composed by several anatomical structures and not only by neurons generating the respiratory rhythm. Certainly, electrophysiological recordings would give us direct information regarding the effect of REM sleep deprivation on VRC neuronal activity. However, for the nature of that technique, it would also have limited us to focus on discrete neuronal populations, and not on all the neuronal populations that form the VRC. An additional limitation for the electrophysiological recording is the depth of the ventral nuclei in the brain stem and the technical difficulty to access them as to perform recording in free movement for prolonged periods.
We conclude that the activity of VRC neurons is highly regulated during sleep to ensure a stable respiratory rate and gas homeostasis necessary to maintain optimum physiology during REM sleep in healthy conditions. We do not rule out direct modulation of anterior structures on motor nuclei.
Beltran-Morgado Foundation for the Advancement and Diffusion of Neuroscience in Veracruz.
6. Conflicto de intereses
The authors declare that there is no conflict of interests.
- Deak MC, Kirsch DB, MD. Sleep-Disordered Breathing in Neurologic Conditions. Clin Chest Med. 2014; 35:547–56.
- Bhat S, Gupta D, Chokroverty S. Sleep Disorders in Neuromuscular Neurol Clin. 2012; 30:1359-87.
- Smith JC, Abdala AP, Rybak IA, Paton JF. Structural and functional architecture of respiratory networks in the mammalian brainstem. Philos Trans R Soc Lond B Biol Sci. 2009; 364:2577–87.
- Alheid GF, McCrimmon DR. The chemical neuroanatomy of breathing. Respir Physiol Neurobiol. 2008; 164:3-11.
- Smith JC, Abdala AP, Koizumi H, Rybak IA, Paton JF. Spatial and functional architecture of the mammalian brain stem respiratory network: a hierarchy of three oscillatory mechanisms. J Neurophysiol. 2007; 98:3370-87.
- Feldman JL, Mitchell GS, Nattie EE. Breathing: rhythmicity, plasticity, chemosensitivity. Annu Rev Neurosci. 2003; 26:239–66.
- Smith JC, Ellenberger HH, Ballanyi K, Richter DW, Feldman JL. Pre-Bötzinger complex: a brainstem region that may generate respiratory rhythm in mammals. Science. 1991; 254:726-9.
- McKay LC, Janczewski WA, Feldman JL. Sleep-disordered breathing after targeted ablation of preBötzinger complex neurons. Nat Neurosci. 2005; 8:1142-44.
- Guyenet PG, Stornetta RL, Bayliss DA. Central respiratory chemoreception. J Comp Neurol. 2010; 518:3883–906.
- Guyenet PG, Stornetta RL, Bayliss DA. Retrotrapezoid nucleus and central chemoreception. J Physiol. 2008; 586:2043-48.
- Grace KP, Horner RL. Evaluating the evidence surrounding pontinecholinergic involvement in REM sleep generation. Front Neurol. 2015; 6:190.
- Szymusiak R, McGinty D. Hypothalamic regulation of sleep and arousal. Ann N Y Acad Sci. 2008; 1129:275–86.
- Snyder F, Hobson JA, Morrison DF, Goldfrank F. Changes in respiration, heart rate, and systolic blood pressure in human sleep. J Appl Physiol. 1964; 19:417–22.
- Mendelson WB, Martin JV, Perlis M, Giesen H, Wagner R, Rapoport SI, Periodic cessation of respiratory effort during sleep in adult rats. Physiol Behav. 1988; 43: 229–34.
- Krieger J, Turlot JC, Mangin P, Kurtz D. Breathing during sleep in normal young and elderly subjects: hypopneas, apneas, and correlated factors. Sleep. 1983; 6:108–20.
- Mendelson WB, Guthrie RD, Frederick G, Wyatt RJ. The flower pot technique of rapid eye movement (REM) sleep deprivation. Pharmacol Biochem Behav. 1974; 2:553–56.
- Paxinos G, Watson C, The Rat Brain in Stereotaxic Coordinates, 5th Edition, Elsevier Academic Press, San Diego California; 2005.
- McKay LC, Feldman JL. Unilateral ablation of pre-Botzinger complex disrupts breathing during sleep but not wakefulness. Am J Respir Crit Care Med. 2008; 178:89–95.
- Rasband, WS. ImageJ, U. S. National Institutes of Health, Bethesda, Maryland, USA, https://imagej.nih.gov/ij/, 1997-2016.
- Radulovacki M, Trbovic SM, Carley DW. Cardiopulmonary interactions following REM sleep deprivation in Sprague-Dawley rats. Exp Neurol. 1997;145(2 Pt 1):371-5.
- Shiromani PJ, Malik M, Winston S, McCarley RW. Time course of Fos-like immunoreactivity associated with cholinergically induced REM sleep. J Neurosci. 1995; 15:3500-8.
- Cirelli C, Pompeiano M, Tononi G. Sleep deprivation and c-fos expression in the rat brain. J Sleep Res. 1995; 4:92-106.
- García-García F, Beltrán-Parrazal L, Jiménez-Anguiano A, Vega-González A, Drucker-Colín R. Manipulations during forced wakefulness have differential impact on sleep architecture, EEG power spectrum, and Fos induction. Brain Res Bull. 1998; 47:317-24.
- Morgan JI, Cohen DR, Hempstead JL, Curran T. Mapping patterns of c-fos expression in the central nervous system after seizure. Science. 1987; 237:192-7.
- Semba K1, Pastorius J, Wilkinson M, Rusak B. Sleep deprivation-induced c-fos and junB expression in the rat brain: effects of duration and timing. Behav Brain Res. 2001; 120: 75–86.
- Jeddi S, Asl AN, Asgari A, Ghasemi A. The Effect of Sleep Deprivation on Cardiac Function and Tolerance to Ischemia-Reperfusion Injury in Male Rats. Arq Bras Cardiol. 2016; 106:41-8.
- Kovalzon VM, Tsibulsky VL. REM-sleep deprivation, stress and emotional behavior in rats. Behav Brain Res. 1984; 14:235-245.
- Rechtschaffen A, Bergmann BM. Sleep deprivation in the rat by the disk-over-water method. Behav Brain Res.1995; 69:55-63.
- Guzman-Marin R., Suntsova N, Bashir T, Nienhuis R. Szymusiak R, McGinty D, Rapid eye movement sleep deprivation contributes to reduction of neurogenesis in the hippocampal dentate gyrus of the adult rat. Sleep. 2008; 31:167-75.
- Gip P, Hagiwara G, Sapolsky RM, Cao VH, Heller HC, Ruby NF. Glucocorticoids influence brain glycogen levels during sleep deprivation. Am J Physiol Regul Integr Comp Physiol. 2004; 286(6):R1057-62.
- Mueller AD, Pollock MS, Lieblich SE, Epp JR, Galea LA, Mistlberger RE. Sleep deprivation can inhibit adult hippocampal neurogenesis independent of adrenal stress hormones. Am J Physiol Regul Integr Comp Physiol. 2008; 294(5):R1693-703.
- Guzmán-Marín R, Suntsova N, Stewart DR, Gong H, Szymusiak R, McGinty D. Sleep deprivation reduces proliferation of cells in the dentate gyrus of the hippocampus in rats. J Physiol. 2003; 549(Pt 2):563-71.
- Joseph V, Pequignot JM, Van Reeth O. Neurochemical perspectives on the control of breathing during sleep. Respir Physiol Neurobiol. 2002; 130:253- 263.
- Orem J. Medullary respiratory neuron activity: relationship to tonic and phasic REM sleep. J Appl Physiol Respir Environ Exerc Physiol. 1980; 48: 54-65.
- Guyanet PG and Mulkey DK. Retrotrapezoid nucleus and parafacial respiratory group. Respir Physiol Neurobiol. 2010; 173(3):244–55.
- Burke PG, Kanbar R, Basting TM, Hodges WM, Viar KE, Stornetta RL, Guyenet PG. State-dependent control of breathing by the retrotrapezoid nucleus. J Physiol. 2015; 593:2909-26.
- Cohen MI, Shaw CF. Role in the inspiratory off-switch of vagal inputs to rostral pontine inspiratory-modulated neurons. Respir Physiol Neurobiol. 2004; 143:127–140.
| Recibido: 10 de julio de 2017 | Aceptado: 08 de octubre de 2017 |
Consuelo Morgado-Valle, Ph.D. Centro de Investigaciones Cerebrales, Universidad Veracruzana. Berlín 7. Fracc. Monte Magno Ánimas. C.P. 91190 Xalapa, Ver. Mexico. Phone: +52(228) 8418900 Ext 16303. E-mail: comorgado@uv.mx
Este es un artículo de libre acceso distribuido bajo los términos de la licencia de Creative Commons, (http://creativecommons.org/licenses/by-nc/3.0), que permite el uso no comercial, distribución y reproducción en algún medio, siempre que la obra original sea debidamente citada.