El papel de los estrógenos y sus receptores en la prevención y promoción de enfermedades proliferativas de la glándula prostática
- Inicio
- Comité Editorial
- Lineamientos
- Carta de Cesión de Derechos
- Información Legal
- Acerca de la Revista
- Bases de Datos
- Contacto
- ISSN 2007-3054
- Centro de Investigaciones Cerebrales
Universidad Veracruzana
Artículo de Revisión
José Locia Espinoza1,2,3* María Elena Hernández Aguilar3 Gonzalo Emiliano Aranda Abreu3 Fausto Rojas Durán3 Jorge Manzo Denes3 Genaro Alfonso Coria Ávila3 Clara Elena Yerena Aguilar1 Abraham Heriberto Soto Cid1,4
1Facultad de Química Farmacéutica Biológica, Universidad Veracruzana, Xalapa, Ver., México 2Doctorado en Neuroetología, Universidad Veracruzana, Xalapa, Ver., México 3Centro de Investigaciones Cerebrales, Universidad Veracruzana, Xalapa, Ver., México 4Instituto Mexicano del Seguro Social, HGZ No. 11.
Resumen/Abstract
Contenido
Conclusiones
Agradecimientos
Referencias
Correspondencia
Los estrógenos son hormonas esteroides que actúan a través de sus receptores (RE) α y β, los cuales actúan como factores de transcripción. A pesar de estar implicados principalmente en la fisiología femenina, los estrógenos son importantes en procesos fisiológicos de los machos, existiendo órganos tales como la próstata cuya homeostasis está influenciada por estas hormonas. En próstata, los estrógenos promueven o limitan la proliferación celular dependiendo del receptor activado, lo cual tiene implicaciones en la fisiopatología de la hiperplasia prostática benigna (HPB) y el cáncer prostático (CaP). Existe evidencia de que RE α promueve proliferación mientras que REβ la regula, comportándose como un factor protector de las patologías mencionadas. No obstante, en etapas avanzadas del CaP, la presencia de REβ en células prostáticas se relaciona con mayor proliferación de éstas y aumento en la mortalidad de los pacientes. Es probable que en este caso REβ active vías de señalización andrógeno-independientes que favorecen la proliferación, lo que resultaría importante en la falta de respuesta al tratamiento de supresión androgénica observado frecuentemente en el CaP. Otro aspecto importante sobre REβ es la regulación de su expresión. En este proceso se conoce la participación de las hormonas esteroides, pero existen indicios de que prolactina, una hormona proteica, participa también en contextos fisiológicos y patológicos. El conocimiento de estos y otros aspectos de la acción estrogénica en próstata contribuirá al desarrollo de estrategias para el tratamiento y prevención de HPB y CaP.
Palabras clave: Estrógenos, Receptores a estrógenos, Próstata, Cáncer de próstata, Prolactina.
Estrogens are steroid hormones acting through their receptors (ER) α and β, act as transcription factors. Despite being primarly involved in female physiology, estrogens are important regulators of some males physiological processes, existing organs such as the prostate, which is influenced by these hormones. In prostate, through its α and β RE estrogens promote or limit the cell proliferation in normal conditions, and are implicated in the pathophysiology of benign prostatic hyperplasia (BPH) and prostate cancer (PCa). There is evidence that REα promotes proliferation, while REβ activation induces protection against the mentioned diseases. However, in advanced stages of prostate cancer, the presence of REβ in prostate cells is associated with increased cell proliferation and increased patient mortality. It is likely that in this case REβ activate signaling pathways that promote androgen-independent proliferation, which would be important in the lack of response to androgen deprivation therapy frequently observed in PCa. REβ another important aspect is the regulation of the ERβ expression. This process is known to involve steroid hormones, but on the other hand, prolactin, a protein pituitary hormone, is also involved in both physiological and pathophysiological contexts. The knowledge of the normal and pathological mechanisms through the estrogens and its β receptors and prolactin regulate the prostate will contribute to the development of strategies for the treatment and prevention of BPH and PCa.
Key words: Estrogens, Estrogen receptors, Prostate, Prostate cancer, Prolactin.
1. Los estrógenos y sus funciones
Los estrógenos son hormonas esteroides con naturaleza no polar, por lo que son capaces de difundir dentro y fuera de las células atravesando la membrana celular con relativa facilidad.1 Debido a ello, los estrógenos actúan a través de receptores intracelulares (los receptores a estrógenos, RE) que son factores de transcripción activados por unión a ligando, es decir son ligando-inducibles.2
Antes del descubrimiento del receptor a estrógenos β (REβ), un estrógeno se definía como tal por su capacidad de estimular la proliferación celular en el útero, un órgano que contiene altas concentraciones del RE que fue identificado y caracterizado como REα.3,4 Puesto que el descubrimiento de REβ ha extendido el panorama con respecto a las acciones estrogénicas en muchos y muy diversos tejidos tanto en machos como en hembras, en la actualidad resulta adecuado clasificar como estrógenos a todas aquéllas moléculas que muestren actividad agonista sobre cualquiera de los dos RE que se conocen.5
En hembras, existen tres estrógenos principales, estos son estradiol (E2), estrona y estriol. De ellos, el E2 es el más abundante, siendo su fuente principal el ovario, mismo que también produce pequeñas cantidades de estrona (Figura 1).
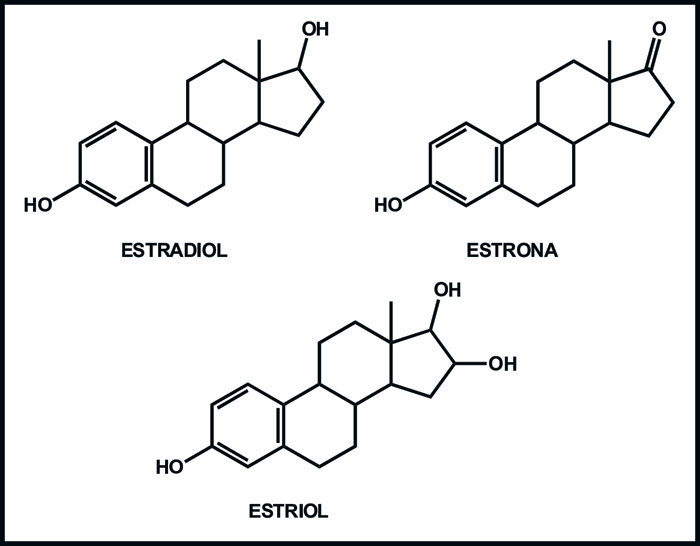
Figura 1. Estructuras de los principales estrógenos fisiológicos.
La fuente principal de esta última son los tejidos periféricos, en tanto que el estriol se produce principalmente en el hígado.6 Los estrógenos también se producen en cantidades menores en glándulas adrenales y tejido adiposo.7,8
Por los efectos que ejercen sobre las funciones reproductivas de la hembra, los estrógenos son considerados principalmente hormonas feminizantes. Entre sus efectos se encuentran los cambios en los depósitos de grasa corporal, el desarrollo y crecimiento mamario, la estimulación de la proliferación celular del útero, trompas de Falopio y epitelio vaginal, la maduración de los ovarios y la producción de hormonas esteroides durante la fase luteal del embarazo.6,9 Asimismo, también se encuentran implicados en el control de la conducta sexual femenina a nivel del sistema nervioso central.8 Por otro lado, además de participar en la función reproductiva, los estrógenos son necesarios para el mantenimiento del equilibrio hidroelectrolítico y de la masa ósea (de ahí los problemas de osteoporosis que se presentan después de la menopausia) y participan en aspectos relacionados con la fisiología vascular tales como la proliferación epitelial de los vasos sanguíneos - siendo factores cardioprotectores - entre otras funciones.2,6
No obstante su importante papel en la fisiología femenina, los estrógenos también son necesarios en procesos fisiológicos de los machos, lo que ha dejado atrás la idea de que las características definitorias de machos y hembras se deben a andrógenos y estrógenos respectivamente. Uno de los efectos estrogénicos más estudiados en el macho es el feedback negativo sobre la secreción de T. Este fenómeno consiste en la inhibición que los estrógenos ejercen sobre la secreción de la hormona luteinizante (LH) en pituitaria.10,11 Puesto que esta hormona se encarga de estimular la secreción de T por parte de las células de Leydig testiculares, el feedback negativo producido por los estrógenos resulta en disminución de los niveles T, que a su vez lleva a una menor producción de DHT.11,12 Por su parte, la ejecución de la conducta sexual y pautas conductuales como la erección y monta requieren de la presencia tanto de andrógenos como de estrógenos a nivel del sistema nervioso central,13,15 donde se ha encontrado expresión alta de RE.16
El desarrollo y función testicular también requieren de los estrógenos, lo cual queda evidenciado por el macroorquidismo y la oligozoospermia que se observan en hombres con niveles bajos de E2. Sin embargo, quizá el efecto estrogénico más importante a nivel del éxito reproductivo del macho sea el control de la reabsorción de fluidos a nivel del epidídimo, proceso que causa un aumento en la cantidad de espermatozoides en el semen y por lo tanto aumenta la probabilidad de que se produzca la fecundación.12,17
Sin duda, otro sitio de acción importante de los estrógenos en el macho es la glándula prostática, cuyo desarrollo y funcionamiento se ven influenciados por acciones estrogénicas tanto indirectas, mediante el control de la producción de andrógenos por feedback negativo explicado anteriormente; como directa; a través de la activación de los RE, principalmente REβ,18-20 que es el objetivo más importante de la presente revisión y se tratará más detalladamente.
En los machos, la principal fuente de estrógenos son los testículos, en cuyas células de Leydig la T se biotransforma en E2 mediante una reacción de aromatización catalizada por enzimas del complejo de citocromos p-450 (CYP450) denominadas aromatasas.21 El E2 así obtenido se vierte a la circulación sistémica y a través de ella alcanza sus tejidos blanco, donde interactúa de manera específica con los RE. También existe la posibilidad de que en el macho tenga mayor importancia la producción tisular local de estrógenos a partir de la T generada en células de Leydig y adipocitos. Lo anterior resulta posible en vista de que las aromatasas se expresan en cantidades significativas en tejidos sensibles a estrógenos.12,16,22
2. El papel de los receptores a estrógenos en la señalización estrogénica
La mayor parte de los efectos de los estrógenos en sus órganos blanco se deben a la cadena de eventos celulares conocida como “señalización estrogénica”, la cual es una vía de transducción celular que inicia con la activación de los RE (Figura 2).
Estos receptores son factores de transcripción que se clasifican dentro de la clase II de la Familia de Receptores Nucleares a hormonas esteroides,22 cuyos integrantes poseen una estructura terciaria que incluye cinco dominios funcionalmente distintos designados con las letras “A” a “F”23 (Figura 3), de éstos, el dominio C es la parte del receptor que se une al ADN (Dominio de Unión al ADN, DUA) en los elementos de respuesta de sus genes blanco, en tanto que el E corresponde al Dominio de Unión a Ligando (DUL) y es el sitio de reconocimiento y unión a la hormona específica y otras moléculas afines.23 En ausencia de ligando, los RE permanecen inactivos, ya que se encuentran enlazados a proteínas chaperonas del complejo HSP (Heath Shock Proteins, proteínas de choque térmico), quienes los mantienen en una conformación que oculta sus péptidos señal de localización nuclear (SLN). La unión del receptor con la hormona correspondiente provoca cambios de conformación que lo separan de las HSP y descubren las señales de localización nuclear; posibilitando su traslocación a núcleo y la activación de la transcripción de genes blanco por interacción con sus elementos de respuesta,24 denominados ERE (Elemento de Respuesta a Estrógenos). Los ERE son secuencias palindrómicas con la estructura 5’-GGTCAnnnTGACC-3’, donde “n” representa un nucleótido cualquiera.25 Aquéllos genes que cuentan con un ERE en su estructura, son clasificados como “responsivos a estrógenos”. Sin embargo, existen genes que aun careciendo de ERE, pueden transcribirse mediante señalización estrogénica gracias a un mecanismo indirecto. En éste, los RE interactúan con otros factores de transcripción tales como Jun, Fos, AP-1 y SP-1, provocando su activación y posterior traslocación a núcleo, donde a su vez se unen con sus propios elementos de respuesta y provocan la expresión génica (Figura 2).22,26,27 La interacción con otros factores de transcripción es diferente dependiendo del tipo de RE implicado, lo que se ha propuesto como explicación al hecho de que la activación de los RE por los mismos ligandos puede provocar efectos distintos dependiendo del contexto tisular y celular. En efecto, una de las características más interesantes de la señalización estrogénica es la existencia de efectos que muchas veces son antagónicos. Por ejemplo, el tratamiento con estrógenos puede resultar en incremento o disminución de la proliferación celular epitelial en útero y próstata,28 de la expresión de apolipoproteínas en hipocampo29 o de la transcripción génica en hueso e hígado30 dependiendo de si el receptor activado es REα o REβ. Las características de estas moléculas serán descritas a continuación.
3. Dos tipos de receptores a estrógenos: REα y REβ
En 1958, se descubrió que las acciones de los estrógenos requerían de la activación de un receptor intracelular, cuya composición y estructura fueron determinadas en 1986 por Green y colaboradores.4 Durante mucho tiempo se pensó que todas las acciones de los estrógenos estaban mediadas por este RE. Sin embargo, en 1993 se generó un ratón transgénico que carecía de su expresión. Contrario a lo que se pensaba, este ratón transgénico (conocido como “ratón ERKO”, por sus siglas en inglés: Estrogen Receptor Knock Out), seguía mostrando respuesta a la administración de E2. Posteriormente, en 1994 se reportó la existencia de un paciente humano que tampoco expresaba el receptor y, aunque padecía osteoporosis e infertilidad, su condición no era incompatible con la vida. Lo anterior - aunado a los mencionados efectos ambiguos observados en respuesta a la administración de estrógenos - hizo pensar en la existencia de más de un receptor para el estradiol y los estrógenos en general. Esta suposición fue confirmada por Kuiper y colaboradores en 1996,18 quienes reportaron el descubrimiento de una segunda molécula capaz de activarse específicamente por interacción con E2 y otros estrógenos
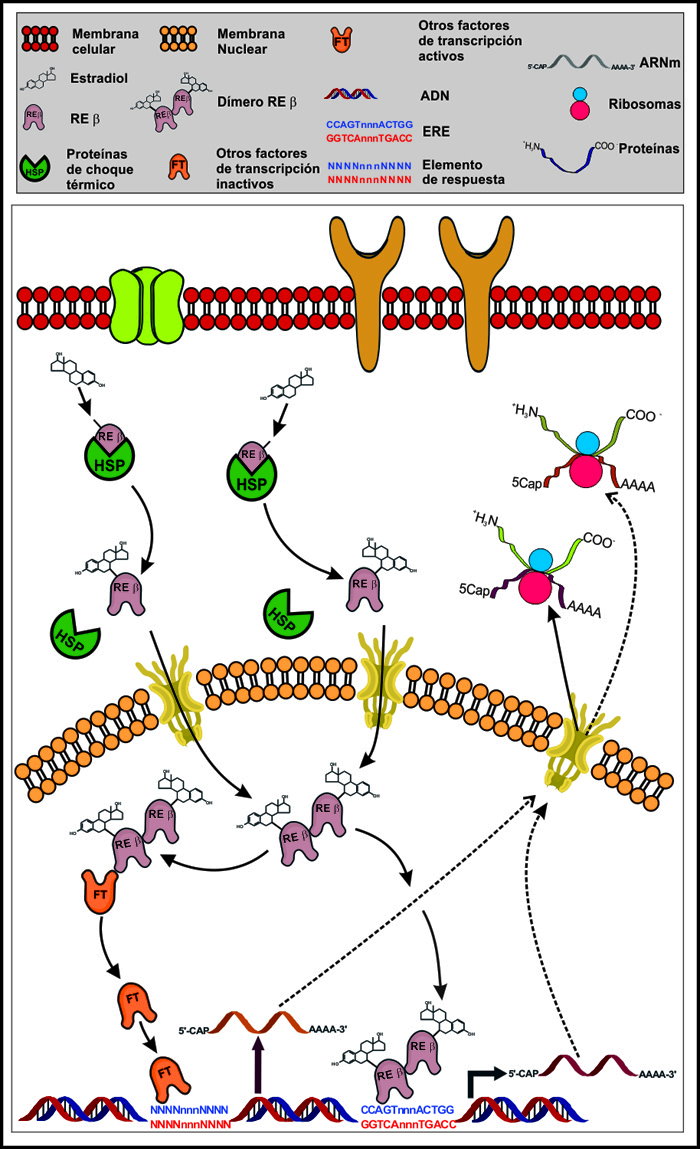
Figura 2. Señalización estrogénica. Los efectos de los estrógenos requieren de la activación de los RE y la consecuente estimulación de la transcripción de genes blanco por interacción directa con el ERE. Los RE también pueden provocar que otros factores de transcripción se unan a sus propios elementos de respuesta.
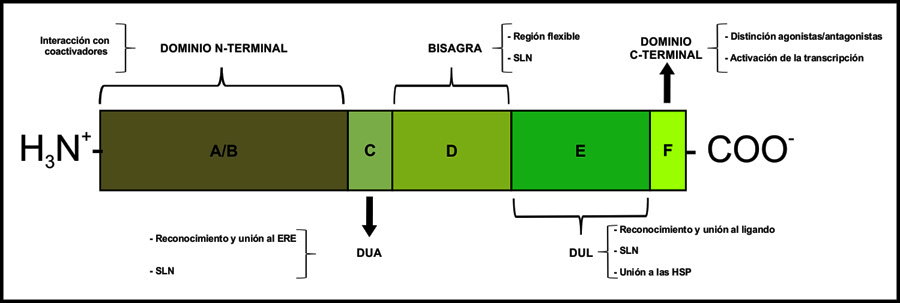
Figura 3. Representación de la estructura terciaria de los receptores a hormonas esteroides. Se mencionan brevemente las funciones de cada dominio. Las abreviaturas se indican en el texto.
El nuevo RE fue identificado en próstata y ovario de rata y, a partir de entonces, el primer RE fue denominado REα, mientras que el segundo es conocido como REβ. Ambas proteínas muestran varias diferencias. Empezando con la longitud y el peso, se sabe que REα tiene una longitud de aproximadamente 600 aminoácidos, mientras que REβ tiene alrededor de 500 aminoácidos. Por lo tanto el peso molecular reportado para cada uno de ellos es 67 y 55 kDa, respectivamente. Con respecto a la secuencia de aminoácidos, se ha mostrado una homología global del 47% entre ambos RE. El dominio que muestra la mayor similitud secuencial en ambos receptores es el C (97%). Puesto que este dominio corresponde al DUA, es decir, la parte de la molécula que interactúa con el ADN para estimular la transcripción, es factible suponer que ambas moléculas se unan al ADN con afinidad y especificidad similares. Sin embargo, cómo se ha mencionado anteriormente, existen efectos particulares que resultan de la activación de uno u otro receptor. Este fenómeno aún no cuenta con una explicación completamente satisfactoria, aunque se han propuesto algunas teorías al respecto. En primer lugar y como ya se ha mencionado, los RE pueden interactuar de forma diferencial con otros factores de transcripción, activándolos y promoviendo así la transcripción de un grupo de genes que no son originalmente sus blancos. Este tipo de acción se denomina comunicación cruzada (o “crosstalk”, en inglés) (Figura 2).26,29 Por otra parte, cuando se han unido al ERE, los RE interactúan de manera también diferencial con moléculas coactivadoras y correpresoras, mismas que tienen un papel importante en la transcripción al producir cambios a nivel de cromatina y otras proteínas importantes en la expresión de genes.3,22 La interacción con ligandos es otra diferencia entre los RE. Si bien ambos muestran su mayor afinidad por E2, REα es aproximadamente cuatro veces más afín que REβ,31 quien por su parte muestra afinidad especialmente alta (aunque más baja que por E2) por los fitoestrógenos7,31 moléculas presentes en la soya y otros productos naturales1,32 cuyo consumo en la dieta reduce la incidencia de cáncer en glándula mamaria, colon y próstata.33,34 De hecho, se considera que el efecto protector contra cáncer de próstata y otros trastornos con proliferación celular incrementada tales como la hiperplasia prostática benigna (HPB) y el carcinoma prostático (CaP) es consecuencia directa de la activación de REβ, mismo que es considerado un regulador de la proliferación celular epitelial prostática. En contraste, existe evidencia de que los efectos estrogénicos proliferativos en próstata (tanto fisiológicos como patológicos) resultan de la activación de REα.28 A continuación, profundizaremos un poco más acerca de los efectos estrogénicos en la glándula prostática.
4. La próstata como blanco de la acción estrogénica
A través de los RE o mediante el control de la producción de andrógenos, el E2 y otros estrógenos influyen en el desarrollo y la fisiología normal de la próstata. Asimismo, las patologías con proliferación celular aumentada en esta glándula tales como la HPB y el CaP también tienen que ver con los estrógenos, no obstante sean los andrógenos los principales responsables de ellas.
La próstata es una glándula sexual accesoria que se encuentra únicamente en machos.35 Como en todas las glándulas sexuales accesorias (vesículas seminales, glándulas bulbouretrales), la función principal de la próstata es producir secreciones que se integran al fluido seminal. Estas secreciones facilitan el trayecto de los espermatozoides a través del tracto reproductor femenino para alcanzar los óvulos durante la cópula y en consecuencia facilitan la fertilización. Específicamente, las secreciones de las glándulas sexuales accesorias aumentan la movilidad espermática, disminuyen la viscosidad del moco cervical y propician la disolución del coágulo seminal.35-37 La próstata es responsable del aporte de aproximadamente 20 % del volumen total de fluido seminal.38
En el humano, la próstata es la glándula accesoria de mayor tamaño. Tiene forma parecida a la de una nuez y se localiza en la pelvis, por debajo de la vejiga y rodeando a la uretra, cuyo segmento que tiene contacto con la próstata es denominado justamente “uretra prostática”.38,39 En su organización microscópica, la próstata es un conjunto de glándulas tubuloalveolares (también llamadas tubuloalveolos) cuyo epitelio es de tipo glandular exócrino,38 el cual está especializado en la secreción hacia conductos que comunican con el exterior.22 Tanto los tubuloalveolos como los conductos, se encuentran inmersos en una mezcla de fibras de tejido conectivo y músculo liso conocido como estroma,35 en el cuál se ha reportado la síntesis local de E2. El E2 sintetizado en el estroma tiene acciones de tipo parácrino sobre el epitelio glandular.40
En el roedor, el modelo animal más utilizado en las investigaciones sobre la próstata; las características del epitelio y el estroma de la glándula son similares a las del humano (Figura 4); no así la organización macroscópica. En roedores, la próstata se encuentra dividida en tres lóbulos individuales conocidos como ventral (PV), lateral (PL) y dorsal (PD). Por su parte, la próstata humana se encuentra dividida en las cuatro zonas siguientes (Figura 5): rodeando a la uretra prostática se localiza la zona transicional, que corresponde al 5 % del total del tejido prostático. Adyacente a la transicional se observa la zona central, que contiene el 25 % del tejido y rodea al ducto eyaculatorio desde la base de la glándula hasta el vero montanum. Por último, comprendiendo aproximadamente el 70% de la glándula prostática, se encuentra la zona periférica, la cuál es la región que puede palparse durante un examen digitorrectal y donde se genera aproximadamente el 90 % de los cánceres prostáticos.38
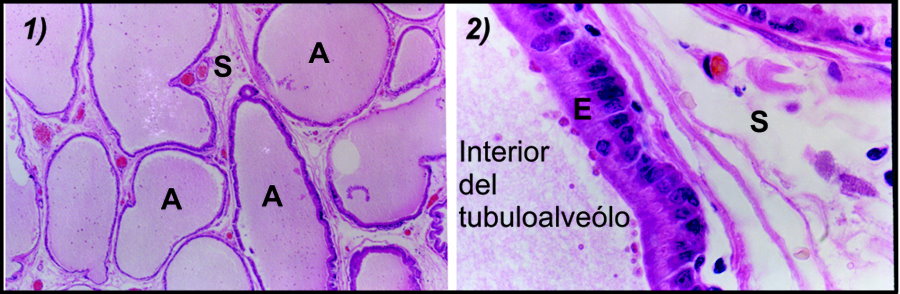
Figura 4. Cortes histológicos de próstata de rata teñidos con hematoxilina y eosina que muestran su organización tubuloalveolar.
Los andrógenos, secretados mayoritariamente por las células de Leydig testiculares son los principales factores hormonales en el control del desarrollo y fisiología de la próstata.35,41,42 En ésta y otros tejidos andrógeno dependientes, es necesario que T, el andrógeno con las mayores concentraciones plasmáticas43 sea metabolizado mediante la enzima 5α-reductasa a DHT, la cual muestra mayor afinidad al receptor a andrógenos (RA),41 por lo tanto, los efectos androgénicos en próstata se deben principalmente a DHT y no a T (Figura 6).
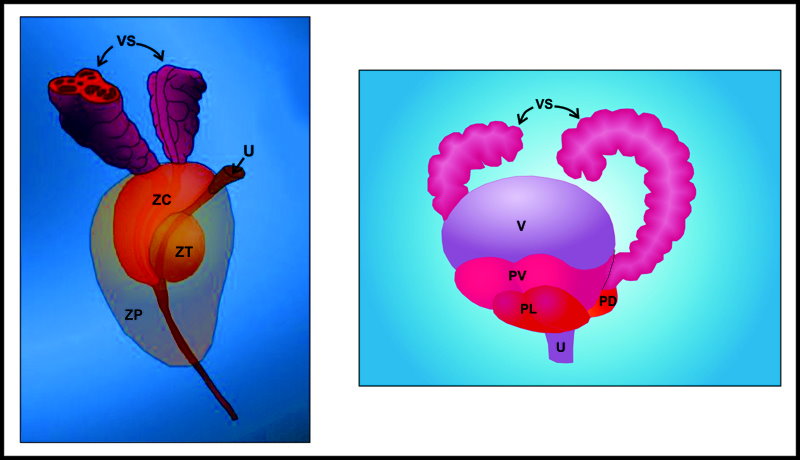
Figura 5. Organización macroscópica de la próstata humana (izquierda) y de rata (derecha).
En lo que respecta al papel de los estrógenos en la glándula prostática, actualmente se acepta que si bien no se les conoce un efecto definido, sí ejercen efectos importantes tanto en condiciones fisiológicas como patológicas. Lo anterior queda demostrado cuando la proporción en plasma de estrógenos/andrógenos se modifica favoreciendo el predominio de los primeros. Existen dos situaciones fisiológicas en las que se presenta este fenómeno. La primera ocurre durante el tercer trimestre de desarrollo fetal, momento en el cual se incrementan los niveles plasmáticos de estrógenos maternos. Este aumento estrogénico está correlacionado con la presencia de metaplasia escamosa en la próstata fetal, que desaparece después del nacimiento, precisamente cuando los estrógenos regresan a sus valores normales. La segunda se observa entre la cuarta y quinta décadas de vida, cuando la producción normal de andrógenos empieza a disminuir, al tiempo que los niveles plasmáticos de proteína fijadora de hormonas sexuales (PFHS) aumentan. Ello ocasiona disminución de T plasmática libre. Asimismo, el incremento en la masa corporal en general y del tejido adiposo en particular posibilitan una mayor expresión de aromatasas tisulares y en consecuencia un aumento en la tasa de conversión de T a E2, lo que trae como consecuencia que la proporción Estrógenos/Andrógenos vuelva a incrementarse y se afecte nuevamente el crecimiento de la próstata, propiciando la aparición de trastornos con proliferación celular aumentada tales como HPB, que tiene su mayor incidencia precisamente en estas etapas de la vida.38,44 De hecho, la inducción de proliferación anormal e hipertrofia celular es una respuesta común a los estrógenos en diversos tejidos, por lo que son considerados carcinógenos potenciales, ya que al aumentar el índice de proliferación también aumentan el índice de replicación del ADN y por ende la probabilidad de que se presenten mutaciones.45
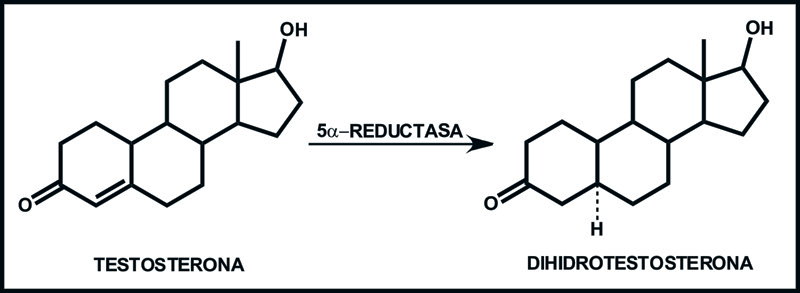
Figura 6. Transformación de T en DHT por la 5-α-reductasa.
La exposición a estrógenos durante la vida gestacional o en el periodo neonatal también tiene consecuencias sobre la proliferación celular prostática en etapas posteriores de la vida. Este fenómeno, que se denomina muy adecuadamente “impronta estrogénica” ha sido ampliamente estudiado en roedores y puede ser un mecanismo fisiológico de programación del tejido glandular para que en etapas posteriores del desarrollo tenga un crecimiento y función determinados.46
Sin embargo, cuando la exposición a estrógenos es excesiva, se correlaciona con susceptibilidad a presentar trastornos hiperproliferativos tanto benignos como malignos. Lo anterior parece ser resultado de cambios en el patrón de expresión de receptores a hormonas esteroides en el epitelio, el cuál pasa de ser un tejido predominantemente andrógeno dependiente a uno con mayor responsividad a estrógenos46,47 y por lo tanto más susceptible al desarrollo de cáncer.48 En humanos, hay indicios de que la estrogenización excesiva durante el desarrollo temprano es igualmente causa de trastornos hiperproliferativos. A este respecto, puede citarse el hecho de que los afroamericanos, quienes manifiestan riesgo elevado de padecer HPB y CaP, también están expuestos a niveles más elevados de estrógenos maternos durante el desarrollo fetal temprano en comparación con los caucásicos.38
Es claro que los estrógenos tienen influencia sobre la glándula prostática, particularmente en lo que respecta al crecimiento y la proliferación celular. En estos procesos, son muy importantes los efectos de los RE, particularmente aquéllos relacionados con REβ, algunos de los cuáles se tratarán en la siguiente sección.
5. El REβ como un regulador de la proliferación celular en próstata
En la próstata el REβ es una proteína característica de las células epiteliales, las cuáles son importantes en la producción de las secreciones de esta glándula. Aunque la presencia de este receptor en el compartimiento epitelial de la glándula prostática ha sido bien caracterizada en numerosas especies, incluyendo al humano, su función no ha sido completamente establecida. Existen diversos estudios que indican que REβ podría estar implicado en el control y regulación de la proliferación glandular, aunque esta hipótesis no es aceptada completamente.
A nivel mundial, el grupo de Jan-Ake Gustafsson del Karolinska Institutet de Suecia es el principal defensor de la teoría que presenta a REβ como un regulador de la proliferación e incluso como un factor de protección contra la hiperplasia y el cáncer. En 1999, este grupo generó un ratón transgénico con fenotipo REβ-/-, es decir, carente de la expresión de REβ (denominado ratón βERKO, en inglés).49 En este organismo se han realizado estudios que parecen confirmar el papel antiproliferativo de REβ. Específicamente, la próstata de los ratones βERKO muestra hiperplasia a partir de los seis meses de edad49 que se acompaña de disminución en la expresión de la enzima glutatión peroxidasa5 y de aumento de la proteína antiapoptótica bcl 2. También se observa incremento en la cantidad de RA, el cual está directamente implicado en la estimulación de la proliferación epitelial prostática,19,50 pues el receptor específico mediante el cual actúan los andrógenos. La próstata de ratones βERKO manifiesta asimismo anomalías en la diferenciación celular, pues la mayoría de las células epiteliales expresan marcadores de células incompletamente diferenciadas tales como p63, y CK 19 en cantidades mayores que en ratones normales.
En próstata humana también se encuentra evidencia de que REβ podría estar controlando la proliferación celular. En primer lugar, la expresión de este receptor es alta en zonas donde la proliferación celular se encuentra restringida. Este es el caso de las células epiteliales luminales en los conductos alveolares, que además cuentan con un grado de diferenciación mayor y por ende participan más en la función secretora de la glándula.7,51 En el estroma, donde existe mayor cantidad de células proliferantes, la expresión de REβ suele ser baja o nula.44,52,53 Resulta importante resaltar que en esta zona prostática existe expresión alta de REα, algo que también se observa en próstata de roedores.18,53-55 Así, es probable que la influencia de la señalización estrogénica en el control de la proliferación y diferenciación celular prostáticas pueda seguir dos rutas con efectos antagónicos. En una de ellas, la activación de REα estromal traería como consecuencia un aumento en el índice de proliferación celular al estimular la producción de factores de crecimiento estromales que tendrían un efecto parácrino sobre las restantes células prostáticas.7,44,56 La segunda ruta iniciaría después de la activación de REβ. Ésta, según se ha argumentado; llevaría al control de la proliferación celular prostática y estimularía la diferenciación.
6. Acciones antagónicas entre REβ REα en la próstata carcinomatosa
El antagonismo que manifiestan los RE en el control de la proliferación y la diferenciación celular tiene implicaciones en el desarrollo, la progresión y el tratamiento del cáncer en diversos tejidos. Como ya se ha expuesto, existe evidencia de que REα promueve, en tanto que REβ limita la proliferación celular, de modo que algunos autores consideran a REβ un factor de protección contra el cáncer, en tanto que REα podría actuar como un oncogen que se sobreexpresa en las células carcinomatosas.7,28 En este sentido, es bastante significativo el hecho de que en algunos cánceres de mama se haya detectado pérdida del cromosoma 14,44 mismo en el que se encuentra localizado el gen codificante de REβ. Por otro lado, cuando se compara la expresión de los receptores en el tejido carcinomatoso con la del tejido sano circundante, suele encontrarse un aumento en la expresión de REα y una disminución en la de REβ en el tejido anormal. Este fenómeno se ha constatado en glándula mamaria,28,57 ovario y colon.28,58 En próstata también existe evidencia de que REβ podría ser un factor de protección contra el cáncer. En principio de cuentas, se tienen los hallazgos hechos en los ratones βERKO que se presentaron en la sección anterior. Además, se ha observado que en especímenes tisulares de próstata humana con diferente grado de proliferación, la cantidad de proteína REβ detectable por inmunohistoquímica (IHQ) desciende conforme aumenta el grado de proliferación, siendo alta en células epiteliales normales, menor en HPB59 y muy baja o ausente en células carcinomatosas. Lo anterior también se acompaña de incremento en la expresión de REα.7,52 Es importante mencionar que el descenso en la expresión de REβ podría ser consecuencia de aumento en el nivel de metilación en islotes CpG del promotor del gen de REβ.44,48,60 Recordemos que la metilación en regiones promotoras de los genes es un mecanismo de inhibición de la expresión génica.61
Los efectos de la activación de los RE que resultan en promoción o inhibición de la proliferación celular no se conocen con exactitud. El REβ podría reducir la proliferación celular a través de la inhibición de la expresión de genes implicados en la replicación del ADN y el ciclo celular.62 Entre estos últimos se encuentra el gen de ciclina D1, factor que controla la transición entre las fases G1 y S del ciclo celular.28,44 El REβ podría inhibir la expresión de estos genes interactuando directamente con sus regiones promotoras e impidiendo su transcripción o bien a través de los ya mencionados mecanismos indirectos en los cuales REβ interactúa con otros factores de transcripción (AP1 y SP1, por ejemplo) y provoca que a su vez éstos estimulen o repriman la expresión de sus propios genes blanco. Lo anterior es posible pues algunos de los genes cuya expresión está controlada por la activación de REβ carecen de EREs en sus promotores.62
Recientemente se ha encontrado que los CaP más agresivos se asocian con la presencia de una traslocación cromosomal adquirida. En esta traslocación, la región promotora del gen de Serina-Proteasa Transmembranal-2 (SPT2) se fusiona con la región codificante de algunos factores de transcripción de la familia de Transformadores Específicos de Eritroblastos (TEE). Los cromosomas con la fusión SPT2-TEE contienen 87 genes que contribuyen a la patología y cuya expresión se ve afectada por la activación de los RE. Específicamente, se ha mostrado que la activación de REα promueve un aumento en la expresión de los genes de SPT2-TTT, mientras que la de REβ promueve una disminución.7
Otro proceso en el cual REβ interviene para regular la proliferación celular prostática tanto en células carcinomatosas como normales es la expresión de RA. Ya hemos mencionado que la proliferación celular en la próstata depende principalmente de los andrógenos, los cuáles requieren de su receptor para ejercer sus efectos a nivel de la activación de la transcripción de genes blanco.35,41 Existen indicios de que una de las funciones de REβ consiste en mantener la expresión de RA en niveles adecuados para evitar una proliferación celular excesiva. Por ejemplo, en ratones βERKO hay cantidades mayores de RA que en ratones normales19,50 y, como era de esperarse un incremento en la proliferación celular.49 Asimismo, el tratamiento de cultivos de células carcinomatosas de próstata humana con ligandos específicos de REβ ocasiona una disminución en la expresión de RA.7 Por su parte, es posible promover incremento en la expresión de RA administrando agonistas específicos de REα,7 lo que constituye otra evidencia del antagonismo entre estos dos receptores.
7. Un enfoque alternativo: el REβ como promotor de carcinogénesis prostática
No obstante la evidencia que liga a las acciones de REβ con un efecto protector contra el cáncer, se ha propuesto que en etapas avanzadas de la enfermedad este receptor podría más bien favorecer el desarrollo de la patología. Un estudio llevado a cabo por Horvath y colaboradores en 2001 con un grupo de 159 muestras provenientes de igual número de pacientes con CaP mostró que la probabilidad de recaída y la mortalidad se encontraba aumentada en pacientes en los que existía expresión de REβ, a diferencia de los que no tenían expresión de este receptor. En el mismo orden de ideas resulta significativo que REβ se exprese en las células DU-145, LNCaP y PC-3.63 Estas células forman parte de líneas estables de carcinoma prostático y, a pesar de encontrarse poco diferenciadas, retienen características de tipo epitelial. Asimismo, salvo LNCaP, no requieren de andrógenos para su crecimiento y proliferación. Por otra parte, fueron establecidas a partir de sitios metastásicos en cerebro (DU-145), nódulos linfáticos (LNCaP) y huero (PC-3).64-66 Así, es posible que REβ tenga un papel bifásico en las patologías prostáticas con proliferación celular aumentada. Al principio, las células normales muestran expresión alta, que se pierde conforme aumenta el nivel de proliferación al grado de que en algunos cánceres desaparece completamente. Esta pérdida de expresión está aparentemente relacionada a incremento en la metilación de islotes CpG en el promotor del receptor.60,67 Posteriormente, REβ vuelve a expresarse en etapas más avanzadas de la enfermedad tales como la metástasis. En ellas, es probable que REβ sea responsable del aumento en la tasa de proliferación celular provocado por estrógenos.60,63,68 Este incremento en la proliferación podría darse a través de la activación de vías de señalización distintas de la estrogénica, como fue demostrado por Pandini y colaboradores en 2007, quienes descubrieron que la expresión del Receptor a Factor de Crecimiento parecido a Insulina (RFCI) se ve incrementada por acción de REβ.69 Este evento es importante puesto que las vías de señalización disparadas por activación de RCFI promueven crecimiento, resistencia a la apoptosis e invasividad de las células cancerígenas.70,71 Así, REβ podría, a través de este mecanismo, favorecer la sobrevivencia de las células carcinomatosas.
Además, el estudio de Pandini y colaboradores (2007) tiene otra implicación importante. Estos investigadores utilizaron cultivos de las líneas celulares que expresan (LNCaP) o no (PC-3) el RA, obteniendo esencialmente los mismos resultados. Esto significa que la proliferación celular en el CaP mediada por los estrógenos puede darse incluso sin la participación de los andrógenos. Esto tiene una relevancia importante desde el punto de vista de que uno de los esquemas de tratamiento contra el CaP consiste en la administración de antagonistas androgénicos con la finalidad de detener la proliferación celular. Esta terapia funciona bien a corto plazo, pero luego se torna insuficiente, dado que la enfermedad se torna insensible a los andrógenos.72 Con lo discutido en este punto, podríamos suponer que parte del mecanismo mediante el cual las células carcinomatosas proliferan en ausencia de andrógenos es la activación de la señalización estrogénica.
8. Mecanismos de regulación de la expresión de REβ en próstata: papel de las hormonas
A lo largo de esta revisión, hemos hecho énfasis en la importancia de la acción hormonal tanto en condiciones normales como patológicas. Conocemos ya parte del papel que juegan estrógenos y andrógenos mediante la activación de sus receptores específicos o al interferir en la acciones de otros mediadores químicos así como en el curso de vías de señalización distintas a la estrogénica. En este punto de nuestro trabajo, nos enfocaremos en el papel que desempeñan algunas hormonas en los procesos de regulación de la expresión de REβ. Siendo este receptor un factor importante en la protección contra el CaP (o incluso un promotor de su progresión en etapas avanzadas), resulta importante conocer los mecanismos mediante los cuales su concentración en esta glándula aumenta o disminuye y en su caso promoverlos o evitarlos, de tal suerte que sea posible conseguir un beneficio en la terapia contra el cáncer o en su prevención. Es importante mencionar que los estudios que buscan determinar la regulación de la expresión de REβ y su relación con las enfermedades proliferativas prostáticas son uno de los enfoques que actualmente buscan esclarecer la fisiopatología del CaP. Algunos de estos enfoques se describen en la Tabla 1.
Las hormonas mayormente implicadas en la regulación de la expresión de REβ son las esteroideas. Sin embargo, como veremos más adelante, la hormona proteica PRL también juega un papel importante y no completamente reconocido en la dinámica de este proceso.
8.1 Regulación por T
Existe evidencia de que los andrógenos juegan un papel importante en la regulación de la expresión de REβ prostático. En rata, por ejemplo, se ha comprobado una disminución en la expresión de este receptor después de la castración, lo que se relaciona con la disminución en la concentración plasmática de T después de la cirugía. El papel de T en este proceso se confirma cuando a los animales castrados se les administra la hormona, pues esto trae como consecuencia una restauración en la expresión de REβ en la glándula.20,54,73 Por otra parte, se ha analizado la expresión normal de REβ en la próstata de ratas de 0 a 3 meses de edad, relacionándola con las variaciones normales en la concentración de T plasmática y se ha encontrado que existe una correspondencia entre las dos moléculas. Así, los niveles circulantes de T se relacionan con la ontogenia de ARNm de REβ en próstata, observándose la mayor expresión de este receptor alrededor de los 90 días de edad, justo cuando T alcanza también sus concentraciones plasmáticas máximas.54,73 Lo anterior se ve apoyado fuertemente por la existencia de una región localizada entre los nucleótidos -98 y -54 del promotor del gen de REβ de rata que es esencial en la estimulación de la transcripción del gen promovida por T. Cuando esta región se elimina, no se observa el aumento en la expresión de REβ en respuesta a T.74 De este modo, el efecto estimulador de la expresión ejercido por T sobre REβ puede ser explicado a partir de la capacidad que tiene la hormona de promover la transcripción del gen del receptor. La cuestión ahora es, ¿cuál es la utilidad o función de la regulación androgénica de la expresión de REβ?, es decir, ¿de qué sirve que T provoque incremento en las concentraciones prostáticas de REβ?; nuevamente, estudios llevados a cabo en roedores arrojan luz sobre esta cuestión. Se ha mencionado ya en esta revisión que la proliferación celular prostática depende principalmente de los andrógenos. En este proceso, T es transformada por la enzima 5α-reductasa a DHT, molécula que muestra una mayor afinidad por RA y por lo tanto es más eficaz en la promoción de la proliferación celular. A su vez, DHT es metabolizada mediante la enzima 3β-hidroxiesteroidorreductasa (3β-HERD) hacia el metabolito 5α-androstan-3β,17β-diol (3β-diol).5 Este derivado androgénico no muestra afinidad significativa por el RA y por lo tanto no se une a él.75 No obstante, 3β-diol es afín a REβ. Aunque la afinidad con que REβ se une a 3β-diol es en términos generales menor que la mostrada por E2,18,31 la concentración prostática del primero es 100 veces más alta que la de este último,19 por lo que es razonable suponer que 3β-diol es un ligando importante de REβ en la glándula prostática. Tomando en cuenta que al principio de esta revisión se definió a un estrógeno por su capacidad de unirse a los RE, 3β-diol puede ser considerado una molécula estrogénica de origen fisiológico específica para REβ. La evidencia de que 3β-diol es una molécula con efectos estrogénicos que suceden por activación de REβ no se encuentra únicamente en los estudios de su afinidad o abundancia en próstata, pues se sabe que su administración en ratas y ratones normales reduce la expresión del RA prostático,19 lo que se acompaña de un descenso en la proliferación celular epitelial,5 efecto que no se observa en ratones βERKO.19 Por otro lado, en ratones que carecen de la enzima 3β-diolhidroxilasa (3β-dH) encargada del metabolismo de 3β-diol, también se observa disminución en la proliferación celular epitelial, ya que al no existir esta enzima, aumentan los efectos antiproliferativos del 3β-diol.5,22 Así, es probable que la presencia de 3β-diol sirva en parte para controlar o limitar la proliferación celular a través de la activación de REβ. De hecho, la producción de este metabolito se ha considerado parte de una vía endócrina relacionada con la proliferación celular en tejido prostático. En esta vía, la enzima 5α-reductasa actuaría sobre T para generar DHT, un andrógeno más potente que estimularía la proliferación celular al activar a RA. Por otra parte, la enzima 3β-HERD actuaría sobre DHT para generar 3β-diol, mismo que al activar a REβ, su receptor específico en próstata; estaría limitando la proliferación celular a niveles aceptables.5,44,76 De esta manera, T estaría mostrando una acción bifásica en cuanto a sus efectos sobre la proliferación celular prostática. Por una parte, es responsable de promoverla a través de sus acciones (directas o a través de DHT) sobre RA, mientras que por otro lado estimula la expresión de REβ y genera al 3β-diol, mismo que al activar a REβ limita la proliferación celular y quizá prevenga el desarrollo de patologías como la HPB y el CaP.
8.2 Regulación por E2
El E2, en términos generales el principal ligando de REβ, también ejerce efectos regulatorios sobre la expresión de éste. Algunos de estos efectos ya se han mencionado en esta revisión cuando se abordó el tema de la “impronta estrogénica”. En este proceso, los efectos no se observan al momento de la exposición a la hormona si no que aparecen en momentos posteriores en la vida de los individuos. La disminución en la expresión de REβ en próstata de ratas es uno de tales efectos.54 En roedores, la inhibición de la expresión de REβ ocasionada por E2 se observa también en pituitaria, testículos,77,78 cerebro y glándula mamaria,79 aunque en estos casos, el efecto aparece al momento de la administración de la hormona. Así pues, los efectos de E2 sobre la expresión de REβ en roedores se observan tanto a corto como a largo plazo. La funcionalidad de esta regulación en próstata y otros tejidos de ratas y ratones no se ha establecido por completo. Es probable que en condiciones normales la hormona contribuya a la regulación de su propio receptor en orden de mantener la homeostasis de los diversos tejidos. Recordemos que E2 no sólo actúa a través de REβ sino que también lo hace a través de REα y que en ambos casos los efectos de la unión son distintos. Asimismo, E2 también tiene influencia sobre la expresión de REα, sólo que en este caso es contraria a la observada para REβ.54,80 De este modo, E2 no sólo actúa de manera antagónica a través de sus receptores sino que también regula su expresión en sentidos contrarios. Así, es probable que bajo ciertas condiciones E2 promueva la expresión de un tipo de receptor mientras que en otras disminuya la del otro tipo con la finalidad de mantener un equilibrio entre las funciones controladas por uno u otro.
Tabla 1. Algunos modelos para el estudio de la relación entre la expresión de REβ y las enfermedades proliferativas prostáticas.
SISTEMA |
OBJETIVO DEL ESTUDIO |
HALLAZGOS IMPORTANTES |
REFERENCIAS |
Ratones |
Efecto de la ausencia de REβ |
HPB y susceptibilidad a CaP. |
|
Rata Wistar |
Efectos de la conducta sexual e hiperprolactinemia. |
Promoción de variaciones en concentraciones séricas de PRL. La hiperprolactinemia ocasiona cambios histológicos que sugieren progresión hacia HPB. |
|
Regulación de la expresión de REβ por T y E2 |
T incrementa expresión de REβ y E2 la disminuye. La estrogenización perinatal ocasiona HPB y susceptibilidad al CaP en individuos adultos. |
||
Humano |
Determinación de la expresión de REβ en muestras histológicas |
Disminución de REβ conforme el tejido progresa hacia la malignidad. Alta en tejido normal, menor en HPB y muy baja o ausente en CaP. |
|
Líneas celulares prostáticas |
Efecto de tratamientos hormonales en la expresión de REβ |
DU-145, PC-3 y LNCaP, células metastásicas, retienen expresión de REβ. La expresión de REβ está controlada por hormonas esteroides y probablemente por PRL. |
8.3 Regulación por PRL
Además de las hormonas esteroides, otros mediadores químicos tienen influencia sobre el crecimiento y funcionalidad de la próstata tanto en contextos fisiológicos como patológicos. Uno de ellos es la hormona proteica prolactina (PRL), cuyos receptores se expresan en esta glándula.81,82 En próstata de roedor, PRL induce cambios en la celularidad de la glándula que indican un incremento en la proliferación celular de la misma.83 Asimismo, en condiciones en las que hay incremento en las concentraciones séricas de PRL, se observa activación de vías de señalización usualmente relacionadas con la unión de PRL a su receptor. Este es el caso de la activación de las proteínas Stat 1 y 3 en próstata durante la ejecución de la conducta sexual en ratas.84 Aunado a lo anterior, es posible que PRL provoque cambios también en la expresión de REβ en la glándula prostática. Ello es posible dada la evidencia que se ha obtenido acerca de este proceso en otras glándulas y tejidos. Así, se observa incremento en la expresión de REβ en cuerpo lúteo,9 pituitaria,85 células deciduales86 y glándula mamaria de rata,79 lo que tiene como consecuencia efectos benéficos sobre la lactancia y la gestación. Se ha propuesto que la activación por PRL de la vía de señalización Jak/Stat (una vía activada por diversas citoquinas que actúan a través de un receptor membranal) es la responsable de promover la estimulación de la expresión de REβ a nivel de la transcripción de su gen (Figura 7).87,88 Es importante mencionar que en esta vía de señalización, pueden tomar parte diversas isoformas de proteínas STAT, algunas de las cuales ya se mencionaron como factores que se activan cuando las concentraciones plasmáticas de PRL son elevadas.84
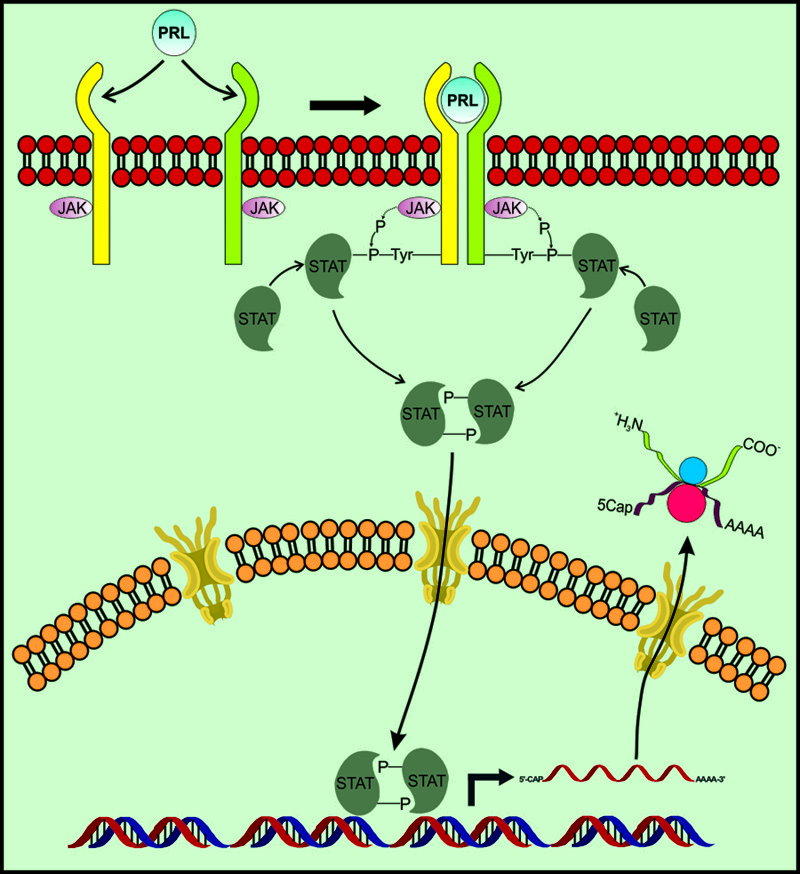
Figura 7. Activación de la vía de señalización Jak/Stat por unión de PRL a su receptor membranal. La unión hormona-receptor provoca dimerización de éste y su posterior fosforilación en tirosinas por una proteína Jak asociada. Las tirosinas fosforiladas se unen a proteínas Stat, que son a su vez fosforiladas por las Jak y después liberadas para dimerizarse y traslocarse a núcleo, donde estimulan la transcripción de sus genes blanco.
Tomando en cuenta lo anterior, es posible que PRL, a través de la activación de sus vías de señalización, tenga influencia sobre la expresión del REβ prostático y que este fenómeno influya positiva o negativamente en la fisiología de la glándula. Esta posibilidad está apoyada por datos aun no publicados de nuestro grupo de trabajo, en los cuales se muestra que la expresión de REβ en próstata de rata incrementa después de que el animal alcanza dos eyaculaciones consecutivas. Este incremento es significativo puesto que ya se ha demostrado que después de dos eyaculaciones las concentraciones séricas de PRL se encuentran también en un máximo.83 Parece probable que este incremento en REβ tiene algo que ver con el aumento en la cantidad de secreciones producidas por la próstata en orden de incrementar el éxito reproductivo. Estudios en los que se observa estimulación de la actividad secretoria de la próstata como resultado del tratamiento con ligandos específicos de REβ tales como el 3β-diol20,38 y el difenilpropionitrilo (DPN)89 respaldan nuestra suposición de que durante la conducta sexual, la estimulación de la expresión de REβ podría tener influencia sobre las secreciones prostáticas.
Por otra parte y siguiendo en línea con nuestras investigaciones, hemos encontrado indicios de que la regulación prostática de REβ mediada por PRL, también se observa durante el cáncer, pues la expresión del mencionado receptor en cultivos de células LNCaP se ve incrementada como resultado del tratamiento con PRL. Previamente hemos discutido el papel deletéreo que REβ podría tener en etapas avanzadas del cáncer de próstata. Por ello, es probable que en estas condiciones la regulación por PRL de la expresión de REβ sea más bien en detrimento del estado de salud del paciente. Sobra decir que es necesario realizar más estudios que confirmen o rechacen nuestras suposiciones.
Los estrógenos cumplen funciones importantes en la fisiología de machos y hembras. También están implicados en el desarrollo y progresión de enfermedades que cursan con proliferación celular incrementada tales como la HPB y el CaP. En estos procesos, es muy importante el papel que juegan sus receptores, los cuales son factores de transcripción capaces de activar no sólo sus genes blanco al interactuar con los elementos de respuesta específicos en los promotores, sino también otros procesos que culminan con un comportamiento específico de la célula.
El mecanismo de acción de los estrógenos a través de sus receptores tiene un panorama complejo y aun no completamente entendido, que se complica aún más por la existencia de los dos tipos de receptores que a la fecha se han identificado. En esta revisión, nos hemos enfocado en el papel que en la glándula prostática cumple REβ, una molécula con aparente función regulatoria de la proliferación celular pero que sin embargo tiene una función dañina en etapas avanzadas del CaP. Entender el funcionamiento de este receptor, así como los mecanismos que regulan su expresión en próstata nos permitirá contar con más elementos para diseñar líneas de acción en la terapia de esta enfermedad que en nuestros días es una causa importante de mortalidad masculina a nivel mundial. En general el estudio de los estrógenos y su acción a través de sus receptores es un campo que ofrece abundantes retos a la investigación tanto básica como aplicada. En la medida en que éste continúe desarrollándose podremos conocer muchas e interesantes funciones que pueden tener impacto tanto en procesos fisiológicos normales como en aquellos que se encuentran involucrados en la generación, progresión y tratamiento de la hiperplasia y el cáncer.
- Beca CONACyT No. 35249 a José Locia Espinoza, con No. de becario 205767.
- Convenio CONACyT 106531.
- Cuerpos académicos Neuroquímica UV-CA-137 y Química Biomolecular UV-CA-202.
- Nilsson S, Makela S, Treuter E, Tujague M, Thomsen J, Andersson G, Enmark E, Pettersson K, Warner M, Gustafsson JA. Mechanisms of estrogen action. Physiol Rev 2001 81: 1535-65.
- Harris HA. Estrogen receptor-beta: recent lessons from in vivo studies. Mol Endocrinol 2007 21: 1-13.
- Koehler KF, Helguero LA, Haldosen LA, Warner M, Gustafsson JA. Reflections on the discovery and significance of estrogen receptor beta. Endocr Rev 2005 26: 465-78
- Green S, Walter P, Kumar V, Krust A, Bornert JM, Argos P, Chambon P. Human oestrogen receptor cDNA: sequence, expression and homology to v-erb-A. Nature 1986 320: 134-9.
- Weihua Z, Lathe R, Warner M, Gustafsson JA. An endocrine pathway in the prostate, ERbeta, AR, 5alpha-androstane-3beta,17beta-diol, and CYP7B1, regulates prostate growth. Proc Natl Acad Sci U S A 2002 99: 13589-94.
- Sherwood L. Fisiología Humana, de las cálulas a los sistemas. México CENGAGE Learning. 2011 pp 971.
- Bonkhoff H, Berges R. The evolving role of oestrogens and their receptors in the development and progression of prostate cancer. Eur Urol 2009 55: 533-42.
- Freeman ME. Neuroendocrine control of the ovarian cycle of the rat. In: Neill JD (Ed.). Knobil and Neill’s physiology of reproduction. United States of America: Elsevier Academic Press. 2006 pp 2327-2388.
- Telleria CM, Zhong L, Deb S, Srivastava RK, Park KS, Sugino N, Park-Sarge OK, Gibori G. Differential expression of the estrogen receptors alpha and beta in the rat corpus luteum of pregnancy: regulation by prolactin and placental lactogens. Endocrinology 1998 139: 2432-42.
- Sandberg AA. Endocrine control and physiology of the prostate. Prostate 1980 1: 169-84.
- Ricke WA, McPherson SJ, Bianco JJ, Cunha GR, Wang Y, Risbridger GP. Prostatic hormonal carcinogenesis is mediated by in situ estrogen production and estrogen receptor alpha signaling. FASEB J 2008 22: 1512-20.
- Sharpe RM. The roles of oestrogen in the male. Trends Endocrin Met 1998 9: 371-7.
- Scordalakes EM, Imwalle DB, Rissman EF. Oestrogen's masculine side: mediation of mating in male mice. Reproduction 2002 124: 331-8.
- Manzo J, Cruz MR, Hernandez ME, Pacheco P, Sachs BD. Regulation of noncontact erection in rats by gonadal steroids. Horm Behav 1999 35: 264-70.
- Hernandez ME, Soto-Cid A, Aranda-Abreu GE, Diaz R, Rojas F, Garcia LI, Toledo R, Manzo J. A study of the prostate, androgens and sexual activity of male rats. Reprod Biol Endocrin 2007 5: 11.
- Wood RI. Estradiol, but not dihydrotestosterone, in the medial amygdala facilitates male hamster sex behavior. Physiol Behav 1996 59: 833-41.
- Hess RA, Bunick D, Lee KH, Bahr J, Taylor JA, Korach KS, Lubahn DB. A role for oestrogens in the male reproductive system. Nature 1997 390: 509-12.
- Kuiper GG, Enmark E, Pelto-Huikko M, Nilsson S, Gustafsson JA. Cloning of a novel receptor expressed in rat prostate and ovary. Proc Natl Acad Sci U S A 1996 93: 5925-30.
- Weihua Z, Makela S, Andersson LC, Salmi S, Saji S, Webster JI, Jensen EV, Nilsson S, Warner M, Gustafsson JA. A role for estrogen receptor beta in the regulation of growth of the ventral prostate. Proc Natl Acad Sci U S A 2001 98: 6330-5.
- Oliveira AG, Coelho PH, Guedes FD, Mahecha GA, Hess RA, Oliveira CA. 5alpha – Androstane - 3beta,17beta - dio (3beta-diol), an estrogenic metabolite of 5alpha-dihydrotestosterone, is a potent modulator of estrogen receptor ERbeta expression in the ventral prostrate of adult rats. Steroids 2007 72: 914-22.
- Hess RA. Oestrogen in fluid transport in efferent ducts of the male reproductive tract. Rev Reprod 2000 5: 84-92.
- Morani A, Warner M, Gustafsson JA. Biological functions and clinical implications of oestrogen receptors alfa and beta in epithelial tissues. J Intern Med 2008 264: 128-42.
- Beato M, Klug J. Steroid hormone receptors: an update. Hum Reprod Update 2000 6: 225-36.
- Sabbah M, Radanyi C, Redeuilh G, Baulieu EE. The 90 kDa heat-shock protein (hsp90) modulates the binding of the oestrogen receptor to its cognate DNA. Biochem J 1996 314:1 205-13.
- Gruber CJ, Gruber DM, Gruber IM, Wieser F, Huber JC. Anatomy of the estrogen response element. Trends Endocrinol Metab 2004 15: 73-8.
- Paech K, Webb P, Kuiper GG, Nilsson S, Gustafsson J, Kushner PJ, Scanlan TS. Differential ligand activation of estrogen receptors ERalpha and ERbeta at AP1 sites. Science 1997 277: 1508-10.
- Liu MM, Albanese C, Anderson CM, Hilty K, Webb P, Uht RM, Price RH, Jr., Pestell RG, Kushner PJ. Opposing action of estrogen receptors alpha and beta on cyclin D1 gene expression. J Biol Chem 2002 277: 24353-60.
- Bardin A, Boulle N, Lazennec G, Vignon, F, Pujol P. Loss of ERbeta expression as a common step in estrogen-dependent tumor progression. Endocr-Relat Cancer 2004 11: 537-51.
- Wang JM, Irwin RW, Brinton RD. Activation of estrogen receptor alpha increases and estrogen receptor beta decreases apolipoprotein E expression in hippocampus in vitro and in vivo. Proc Natl Acad Sci U S A 2006 103: 16983-8.
- Lindberg MK, Moverare S, Skrtic S, Gao H, Dahlman-Wright K, Gustafsson JA, Ohlsson C. Estrogen receptor (ER)-beta reduces ERalpha-regulated gene transcription, supporting a "ying yang" relationship between ERalpha and ERbeta in mice. Mol Endocrinol 2003 17: 203-8.
- Kuiper GG, Carlsson B, Grandien K, Enmark E, Haggblad J, Nilsson S, Gustafsson JA. Comparison of the ligand binding specificity and transcript tissue distribution of estrogen receptors alpha and beta. Endocrinology 1997 138: 863-70.
- Cassidy A. Potential tissue selectivity of dietary phytoestrogens and estrogens. Curr Opin Lipidol 1999 10: 47-52.
- Wanibuchi H, Seok Kang J, Salim EI, Morimura K, Fukushima S. Toxicity vs. beneficial effects of phytoestrogens. Pure Appl Chem 2003 75, 6.
- Sugiyama N, Barros RP, Warner M, Gustafsson JA. ERbeta: recent understanding of estrogen signaling. Trends Endocrinol Metab 2010 21: 545-52.
- Risbridger GP, Taylor AR. Physiology of the male accesory sex structures: The prostate gland, seminal vesicles, and bulbouretral glands. In: Neill JD (Ed.). Knobil and Neill’s Physiology of Reproduction. United States of America: Elsevier Academic Press. 2006 pp 1149-1172.
- Ruenwongsa P y Chulavatnatol M. Acidic protease from human seminal plasma. Purification and some properties of active enzyme and of proenzyme. J Biol Chem 1975 250: 7574-8.
- Aumuller G y Seitz J. Cytochemistry and biochemistry of acid phosphatases. VI: Immunoelectron microscopic studies on human prostatic and leukocytic acid phosphatases. Prostate 1985 7: 161-9.
- Ahmed MM, Lee TC, Oesterling JE. Current trends in the epidemiology of prostatic diseases: Benign hyperplasia and adenocarcinoma. In: J. NR (Ed.). Prostate: basic and clinical aspects. United States of America: CRC Press. 1997 pp 3-25.
- Ross MH y Pawlina W. Histología. Texto y atlas color con biología celular y molecular. Editorial Médica Panamericana, Argentina. 2007 pp. 808-813.
- McPherson SJ, Ellem SJ, Simpson ER, Patchev V, Fritzemeier KH, Risbridger GP. Essential role for estrogen receptor beta in stromal-epithelial regulation of prostatic hyperplasia. Endocrinology 2007 148: 566-74.
- So AI, Hurtado-Coll A, Gleave ME. Androgens and prostate cancer. World J Urol 2003 21: 325-37.
- Heemers HV, Tindall DJ. Androgen receptor (AR) coregulators: a diversity of functions converging on and regulating the AR transcriptional complex. Endocr Rev 2007 28: 778-808.
- Norman AW, Litwack G. Steroid hormones: Chemistry, biosynthesis, and metabolism. In: Norman AW (Ed.). Hormones. United States of America: Academic Press. 1997 pp 49-86.
- Kawashima H y Nakatani T. Involvement of estrogen receptors in prostatic diseases. Int J Urol 2012 19: 512-22.
- Jefcoate CR, Liehr JG, Santen RJ, Sutter TR, Yager JD, Yue W, Santner SJ, Tekmal R, Demers L, Pauley R, Naftolin F, Mor G, Berstein L. Tissue-specific synthesis and oxidative metabolism of estrogens. J Natl Cancer I 2000 95-112.
- Prins GS, Huang L, Birch L, Pu Y. The role of estrogens in normal and abnormal development of the prostate gland. Ann NY Acad Sci 2006 1089: 1-13.
- Prins GS. Developmental estrogenization of the prostate gland. In: Naz RK (Ed.). Prostate: Basic and Clinical Aspects. United States of America: CRC Press. 1997 pp 245-263.
- Hartman J, Strom A, Gustafsson JK. Current concepts and significance of estrogen receptor beta in prostate cancer. Steroids 2012 77: 1262-1266.
- Krege JH, Hodgin JB, Couse JF, Enmark E, Warner M, Mahler JF, Sar M, Korach KS, Gustafsson JA, Smithies O. Generation and reproductive phenotypes of mice lacking estrogen receptor beta. Proc Natl Acad Sci U S A 1998 95: 15677-82.
- Weihua Z, Warner M, Gustafsson JA. Estrogen receptor beta in the prostate. Mol Cell Endocrinol 2002 193: 1-5.
- Cowley SM, Hoare S, Mosselman S, Parker MG. Estrogen receptors alpha and beta form heterodimers on DNA. J Biol Chem 1997 272: 19858-62.
- Pasquali D, Staibano S, Prezioso D, Franco R, Esposito D, Notaro A, De Rosa G, Bellastella A, Sinisi AA. Estrogen receptor beta expression in human prostate tissue. Mol Cell Endocrinol 2001 178: 47-50.
- Royuela M, de Miguel MP, Bethencourt FR, Sanchez-Chapado M, Fraile B, Arenas MI, Paniagua R. Estrogen receptors alpha and beta in the normal, hyperplastic and carcinomatous human prostate. J Endocrinol 2001 168: 447-54.
- Prins GS, Marmer M, Woodham C, Chang W, Kuiper G, Gustafsson JA, Birch L. Estrogen receptor-beta messenger ribonucleic acid ontogeny in the prostate of normal and neonatally estrogenized rats. Endocrinology 1998 139: 874-83.
- Pelletier G, Labrie C, Labrie F. Localization of oestrogen receptor alpha, oestrogen receptor beta and androgen receptors in the rat reproductive organs. J Endocrinol 2000 165: 359-70.
- Yu L, Wang CY, Shi J, Miao L, Du X, Mayer D, Zhang J. Estrogens promote invasion of prostate cancer cells in a paracrine manner through up-regulation of matrix metalloproteinase 2 in prostatic stromal cells. Endocrinology 2011 152: 773-81.
- Leygue E, Dotzlaw H, Watson PH, Murphy LC. Altered estrogen receptor alpha and beta messenger RNA expression during human breast tumorigenesis. Cancer Res 1998 58: 3197-201.
- Burns KA, Korach KS. Estrogen receptors and human disease: an update. Arch Toxikol 2012 86: 1491-504.
- Asgari M y Morakabati A. Estrogen receptor beta expression in prostate adenocarcinoma. Diagn Pathol 2011 6: 61.
- Takahashi Y, Perkins SN, Hursting SD, Wang TT. 17beta-Estradiol differentially regulates androgen-responsive genes through estrogen receptor-beta- and extracellular-signal regulated kinase-dependent pathways in LNCaP human prostate cancer cells. Mol Carcinogen 2007 46: 117-29.
- Alberts B, Johnson A, Lewis J, Raff M, Roberts K, Walter P. In Alberts B (Ed.). Biología molecular de la célula. Ediciones Omega, España. 2004 pp 375-468.
- Zhao C, Dahlman-Wright K, Gustafsson JA. Estrogen receptor beta: an overview and update. Nucl Recept Signal 2008 6: e003.
- Lau KM, LaSpina M, Long J, Ho SM. Expression of estrogen receptor (ER)-alpha and ER-beta in normal and malignant prostatic epithelial cells: regulation by methylation and involvement in growth regulation. Cancer Res 2000 60: 3175-82.
- Stone KR, Mickey DD, Wunderli H, Mickey GH, Paulson DF. Isolation of a human prostate carcinoma cell line (DU 145). Int J Cancer 1978 21: 274-81.
- Horoszewicz, J.S. et al. Kaighn ME, Narayan KS, Ohnuki Y, Lechner JF, Jones LW. Establishment and characterization of a human prostatic carcinoma cell line (pc-3). Investigative urology. 1979;17:16-23
- Horoszewicz, J.S. et al. LNCaP model of human prostatic carcinoma. Cancer Res 1983 43: 1809-18.
- Zhu X, Leav I, Leung YK, Wu M, Liu Q, Gao Y, McNeal JE, Ho SM. Dynamic regulation of estrogen receptor-beta expression by DNA methylation during prostate cancer development and metastasis. Am J Pathol 2004 164: 2003-12.
- Maggiolini M, Recchia AG, Carpino A, Vivacqua A, Fasanella G, Rago V, Pezzi V, Briand PA, Picard D, Ando S. Oestrogen receptor beta is required for androgen-stimulated proliferation of LNCaP prostate cancer cells. J Mol Endocrinol 2004 32: 777-91.
- Pandini G, Genua M, Frasca F, Squatrito S, Vigneri R, Belfiore A. 17beta-estradiol up-regulates the insulin-like growth factor receptor through a nongenotropic pathway in prostate cancer cells. Cancer Res 2007 67: 8932-41.
- LeRoith D y Roberts CTJr. The insulin-like growth factor system and cancer. Cancer Lett 2003 195: 127-37.
- Baserga R, Peruzzi F, Reiss K. The IGF-1 receptor in cancer biology. Int J Cancer 2003 107: 873-7.
- Schulman C, Irani J, Aapro M. Improving the management of patients with prostate cancer receiving long-term androgen deprivation therapy. BJU Int 2012 6: 13-21.
- Asano K, Maruyama S, Usui T, Fujimoto N. Regulation of estrogen receptor alpha and beta expression by testosterone in the rat prostate gland. Endocr J 2003 50: 281-7.
- Fujimoto N, Asano K, Usui T, Kitamura S. Cloning and characterization of the 5'-flanking region of the rat estrogen receptor beta gene. J Steroid Biochem 2005 94: 15-21.
- Turcotte G, Chapdelaine A, Roberts KD, Chevalier S. Androgen binding as evidenced by a whole cell assay system using cultured canine prostatic epithelial cells. J Steroid Biochem 1988 29: 69-76.
- Al-Nakhle H, Burns PA, Cummings M, Hanby AM, Hughes TA, Satheesha S, Shaaban AM, Smith L, Speirs V. Estrogen receptor {beta}1 expression is regulated by miR-92 in breast cancer. Cancer Res 2010 70: 4778-84.
- Tena-Sempere M, Navarro VM, Mayen A, Bellido C, Sanchez-Criado JE. Regulation of estrogen receptor (ER) isoform messenger RNA expression by different ER ligands in female rat pituitary. Biol Reprod 2004 70: 671-8.
- Tena-Sempere M, Navarro J, Pinilla L, Gonzalez LC, Huhtaniemi I, Aguilar E. Neonatal exposure to estrogen differentially alters estrogen receptor alpha and beta mRNA expression in rat testis during postnatal development. J Endocrinol 2000 165: 345-57.
- Hatsumi T y Yamamuro Y. Downregulation of estrogen receptor gene expression by exogenous 17beta-estradiol in the mammary glands of lactating mice. Exp Biol M 2006 231: 311-6.
- Mitchner NA, Garlick C, Ben-Jonathan N. Cellular distribution and gene regulation of estrogen receptors alpha and beta in the rat pituitary gland. Endocrinology 1998 139: 3976-83.
- Bole-Feysot C, Goffin V, Edery M, Binart N, Kelly PA. Prolactin (PRL) and its receptor: actions, signal transduction pathways and phenotypes observed in PRL receptor knockout mice. Endocr Rev 1998 19: 225-68.
- Ouhtit A, Morel G, Kelly PA. Visualization of gene expression of short and long forms of prolactin receptor in rat reproductive tissues. Biol Reprod 1993 49: 528-36 .
- Hernandez ME, Soto-Cid A, Rojas F, Pascual LI, Aranda-Abreu GE, Toledo R, Garcia LI, Quintanar-Stephano A, Manzo J. Prostate response to prolactin in sexually active male rats. Reprod Biol Endocrin 2006 4: 28.
- Soto-Cid A, Hernandez-Kelly LC, Hernandez ME, Manzo J, Gonzalez-Mejia ME, Zepeda RC, Ortega A. Signal transducers and activators of transcription 1 and 3 in prostate: effect of sexual activity. Life Sci 2006 79: 919-24.
- Khurana S, Ranmal S, Ben-Jonathan N. Exposure of newborn male and female rats to environmental estrogens: delayed and sustained hyperprolactinemia and alterations in estrogen receptor expression. Endocrinology 2000 141: 4512-7.
- Tessier C, Deb S, Prigent-Tessier A, Ferguson-Gottschall S, Gibori GB, Shiu RP, Gibori G. Estrogen receptors alpha and beta in rat decidua cells: cell-specific expression and differential regulation by steroid hormones and prolactin. Endocrinology 2000 141: 3842-51.
- Frasor J, Park K, Byers M, Telleria C, Kitamura T, Yu-Lee LY, Djiane J, Park-Sarge OK, Gibori G. Differential roles for signal transducers and activators of transcription 5a and 5b in PRL stimulation of ERalpha and ERbeta transcription. Mol Endocrinol 2001 15: 2172-81.
- Frasor J, Barkai U, Zhong L, Fazleabas AT, Gibori G. PRL-induced ERalpha gene expression is mediated by Janus kinase 2 (Jak2) while signal transducer and activator of transcription 5b (Stat5b) phosphorylation involves Jak2 and a second tyrosine kinase. Mol Endocrinol 2001 15: 1941-52.
- Taylor RA, Cowin P, Couse JF, Korach KS, Risbridger GP. 17beta-estradiol induces apoptosis in the developing rodent prostate independently of ERalpha or ERbeta. Endocrinology 2006 147: 191-200.
| Recibido: 24 de junio de 2013 | Aceptado: 30 de agosto de 2013 |
Correspondencia:
Correspondencia: Laboratorio de Neuroendocrinología, Centro de Investigaciones Cerebrales, Universidad Veracruzana Av. Dr. Luis Castelazo s/n, Col. Industrial Las Animas Xalapa, Ver. C.P. 91190 Teléfono: (228) 8418900 Ext. 13616 Correo electrónico: jlocia@uv.mx
Este es un artículo de libre acceso distribuido bajo los términos de la licencia de Creative Commons, (http://creativecommons.org/licenses/by-nc/3.0), que permite el uso no comercial, distribución y reproducción en algún medio, siempre que la obra original sea debidamente citada.