El status epilepticus no modifica la memoria de trabajo en ratas de 21 días de edad: un estudio preliminar
- Inicio
- Comité Editorial
- Lineamientos
- Carta de Cesión de Derechos
- Información Legal
- Acerca de la Revista
- Bases de Datos
- Contacto
- ISSN 2007-3054
- Centro de Investigaciones Cerebrales
Universidad Veracruzana
Artículo de Investigación
Jesús Servando Medel-Matus1,4, Rosa Angélica Medel-Matus1, Joel Martínez-Quiroz3, Deysi Y. Bermúdez-Ocaña5, César A. Pérez-Estudillo1,2, Luis I. García1,2, Genaro A. Coria-Ávila1,2, María-Leonor López-Meraz1,2
1Centro de Investigaciones Cerebrales, Universidad Veracruzana. 2Facultad de Medicina, Universidad Veracruzana. 3Facultad de Química Farmacéutica Biológica, Universidad Veracruzana y Escuela Nacional de Ciencias Biológicas, Instituto Politécnico Nacional. 4Doctorado en Neuroetología, Universidad Veracruzana. 5División Académica Multidisciplinaria de Comalcalco, Universidad Juárez Autónoma de Tabasco.
Resumen/Abstract
Introducción
Material y métodos
Resultados
Discusión
Conclusión
Agradecimientos
Referencias
Correspondencia
Introducción.El status epilepticus (SE) es un tipo de actividad epiléptica que se caracteriza por la presencia de crisis convulsivas por un tiempo prolongado, manifestándose con mayor frecuencia en la población pediátrica. Entre las consecuencias del SE en humanos y en animales de experimentación se encuentra el deterioro en los procesos de memoria y cognición. Sin embargo, no existen estudios realizados poco tiempo después de que suceda el SE en animales en desarrollo para evaluar sus efectos en la memoria de trabajo. Objetivos.Determinar si el SE causado en animales de 14 días de edad deteriora la memoria de trabajo al ser evaluado durante el día posnatal 21, utilizando el modelo de reconocimiento de un objeto nuevo. Material y métodos. Se utilizaron ratas de la cepa Wistar de ambos géneros. Las ratas se inyectaron con cloruro de litio (LiCl, 3 mEq/Kg, i.p.) el día postnatal 13 y 20 h después el SE se indujo tras las administración de clorhidrato de pilocarpina (100 mg/Kg, s.c.; n=10). El grupo control se inyectó con LiCl y solución salina (n=8). El día posnatal 21 se analizó la memoria de trabajo con el modelo de reconocimiento de un objeto nuevo. Resultados. La exploración del objeto nuevo y familiar no difirió entre las ratas con SE y el grupo control; tampoco el índice de discriminación contrasto entre los grupos de animales. Conclusión.El SE inducido en animales de 14 días de edad no modifica la memoria de trabajo evaluada durante el día posnatal 21.
Palabras clave: Status epilepticus, memoria de trabajo, ratas en desarrollo, modelo de reconocimiento de un objeto nuevo.
Introduction. Status epilepticus (SE) is a neurological condition characterized by continuous seizures during a prolonged period or by seizures occurring so frequently that an individual does not fully recover from one seizure before having another. SE occurs more frequently in the pediatric population and causes long term deficits in learning and memory in humans and experimental animals. However, no evidence exists about the effect of SE induced in developing rats on working memory. Objective. To study the effect of SE induced in fourteen days old rat pups (P14) on working memory during the twenty-one postnatal day (P21). Methods. SE was induced by using the lithium-pilocarpine model: rats were given 3 mEq/Kg lithium chloride (LiCl) i.p. on the day before the induction of SE, which was carried out at P14 by a subcutaneous injection of 100 mg/Kg pilocarpine hydrochloride. Control animals were given an equal volume of LiCl and saline subcutaneously, respectively. Memory was evaluated by using the novel recognition object model at P21. Results. SE did not modify the time of exploration of the familiar and novel object nor the discrimination index as compared with control group. Conclusion. SE induced in P14 rats does not modify working memory evaluated during the P21.
Palabras clave: Status epilepticus, working memory, developing rats, novel recognition object model.
El SE o estado epiléptico es un tipo de actividad no autolimitada, que es caracterizada por crisis epilépticas continuas por un tiempo prolongado o intermitentes sin que el individuo recupere por completo la consciencia entre éstas.1,2 El SE ocurre con mayor frecuencia en niños que en adultos.3-5 A nivel experimental, el SE experimental produce alteraciones conductuales a largo plazo cuando se induce durante la etapa posnatal temprana de la rata,6,7 pero el efecto es diferente dependiendo de la edad del animal a la cual se presenten las convulsiones.8,9
El SE inducido con ácido kaínico en ratas de 14 días de edad produce deterioro en la memoria y el aprendizaje espacial a corto y largo plazo en la adultez.8,9 Sin embargo, pocos son los trabajos que han explorado las consecuencias del SE causado en la rata en desarrollo sobre los procesos mnemónicos poco después de ocurrido el evento convulsivo y sobre otros tipos de memoria a corto plazo, tal es el caso de la memoria de trabajo. Los datos al respecto son de importancia al considerar que en la clínica el SE es más frecuente en la población pediátrica e infantil.3
La memoria de trabajo es una forma de almacenar información de manera transitoria (memoria a corto plazo), es limitada en capacidad y requiere de ensayo,10 pero es de amplia importancia para llevar a cabo las actividades de la vida cotidiana. Esto nos llevó a preguntarnos si el SE que se causa durante etapas tempranas de la vida modifica únicamente la memoria espacial cuando se es adulto o puedo afectar también a la memoria de trabajo poco después del evento convulsivo. Por lo que el propósito del presente estudio fue evaluar si el SE en ratas de 14 días de edad modifica la memoria de trabajo evaluada durante el día posnatal 21. Para este fin se utilizó el modelo de litio-pilocarpina para generar las convulsiones y el modelo de memoria de reconocimiento de un objeto nuevo.
Sujetos de estudio
Se utilizaron ratas de la cepa Wistar criadas en el bioterio del Centro de Investigaciones Cerebrales de la Universidad Veracruzana. Las madres gestantes, y después las madres con sus crías, se mantuvieron a temperatura ambiente, con libre acceso a agua y alimento y con ciclo normal luz-oscuridad de 12 h. Se utilizaron animales de ambos sexos y el día de nacimiento se consideró como el día posnatal cero (P0). En todos los casos, la inducción del SE se realizó a cabo entre las 0900 y 1000 h del día posnatal 14. Al finalizar el evento epiléptico, las crías se regresaron al nido y permanecieron con sus madres hasta el final de los experimentos. Todos los procedimientos se realizaron bajo la normatividad vigente para el uso y cuidado de animales de investigación de México (NOM-062-ZOO-1999).
Inducción del status epilepticus
Durante el día posnatal 13 las ratas se inyectaron con LiCl (3 mEq/Kg, i.p; Sigma) y después de 20 h, durante el día postnatal 14, se administró clorhidrato de pilocarpina (100 mg/Kg, s.c.; Sigma) para inducir el SE11 (n=10). El grupo de animales control se inyectó únicamente con LiCl y solución salina (n=8). Todos los animales se rehidrataron con solución salina glucosada 5% (1 ml, s.c.) 6 h después de la inducción del SE y se regresaron con sus madres. Se trató de modelar un tipo severo de SE, por lo que las convulsiones no se detuvieron con ningún fármaco antiepiléptico.
La manifestación de la actividad convulsiva se monitoreó al registrar las mioclonias de miembros anteriores y cabeza, crisis generalizadas ya sea con pérdida o no de la postura corporal y sólo aquellos animales que desarrollaron SE fueron incluidos en el estudio. Se determinó la latencia y duración del SE, así como el porcentaje de animales que lo presentaron.
Modelo de reconocimiento de un objeto nuevo
El modelo consistió en someter a la rata a una sesión de entrenamiento y posteriormente retarla para evaluar su capacidad para reconocer un objeto nuevo (memoria de trabajo). Previo al experimento, los animales fueron habituados a la cámara de acrílico (40 X 40 cm) durante 5 min por cuatro sesiones (P16-P19) (Figura 1). Al día siguiente (P20), las ratas se sometieron a una sesión de pre-prueba, durante la cual se expusieron en la misma cámara a dos objetos idénticos (cilindros de plástico de 4.5 cm de diámetro x 3.7 cm de alto) durante 5 min; veinticuatro h después (P21), los animales se colocaron en el mismo compartimento por 5 min, pero en este caso uno de los objetos fue nuevo para las ratas, i.e. no presentado antes (figura en forma de cruz de plástico de 4.7 cm de longitud x 3.7 cm de alto) y el otro fue el mismo de la sesión de entrenamiento (objeto familiar).
Las sesiones se videograbaron y se analizó la exploración que la rata hizo a los objetos tanto familiar como nuevo. La exploración se definió como el olfateo o el contacto que la rata realizó sobre el objeto con la nariz o las extremidades delanteras.12 El tiempo total que cada animal exploró los objetos nuevo y familiar se determinó con el Programa de Observación de Comportamiento, (BOP por sus siglas en inglés; Steve Cabilio, 1998). A partir de los tiempos de exploración registrados se calculó el índice de discriminación (ID) para establecer la preferencia de la rata a la exploración del objeto nuevo.12
Los experimentos se realizaron entre las 1200 y las 1400 h. La cámara y los objetos utilizados en las pruebas se limpiaron con etanol al 70% y se dejaron secar antes de la prueba y entre cada rata para eliminar residuos y olores que pudieran interferir con el estudio.
Análisis estadístico
Se verificó que los datos cumplieran los supuestos de normalidad y homogeneidad de varianzas (para no cometer error tipo II) realizando las pruebas de Kolmogorov-Smirnov y Levene, respectivamente. El tiempo de exploración del objeto nuevo y del objeto familiar, así como los valores de ID del grupo control y del grupo con SE se compararon utilizando una prueba de t de Student y se presentan como la media ± el error estándar de la media (E.E.M.).
tiempo explorando el objeto nuevo – tiempo explorando el objeto familiar x 100
ID = ![]()
tiempo explorando el objeto nuevo + tiempo explorando el objeto familiar
El número de animales que presentaron un ID negativo en cada grupo experimental se comparó utilizando la prueba exacta de Fisher. Los análisis se realizaron con el programa Sigma Stat versión 3.5 y para evaluar la existencia de diferencias estadísticamente significativas entre los grupos experimentales se consideró un nivel alfa de p< 0.05.
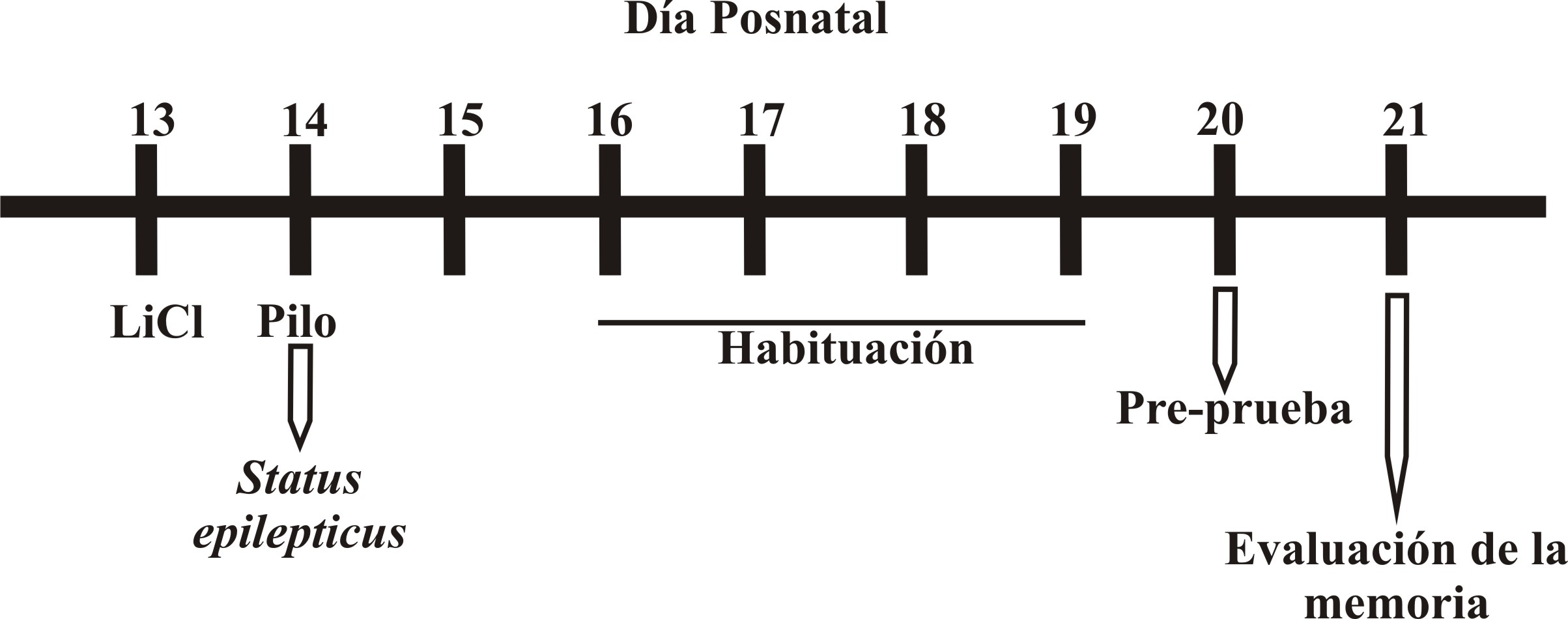
Figura 1. Representación temporal de la inducción del SE, la habituación y evaluación de la memoria de trabajo con el modelo de reconocimiento de un objeto nuevo.
Inducción del SE
El total de los animales tratados con pilocarpina presentaron SE con un tiempo de latencia de 9.3 ± 0.5 min y una duración de 4.7 ± 0.03 h. Las conductas que se observaron en todos los animales fueron en orden de aparición: temblor, mioclonias de cabeza, mioclonias de miembros anteriores y crisis generalizadas caracterizadas por las conductas antes mencionadas y el erguimiento del animal en sus extremidades traseras con pérdida de la postura.
SE y memoria de trabajo
El tiempo de exploración del objeto nuevo (13.3 ± 2.7 s, 57% del tiempo total de exploración) y el objeto familiar (11.7 ± 1.7s, 46% del tiempo total de exploración) por las ratas que presentaron SE fue similar al de los sujetos control [13.3 ± 2.6 s (57% del tiempo total de exploración) y 12.0 ± 2.5 s (43% del tiempo total de exploración), respectivamente] (objeto nuevo: t=0.123, gl=16, p=0.990; objeto familiar t=0.898, gl=16, p=0.930) (figura 2).
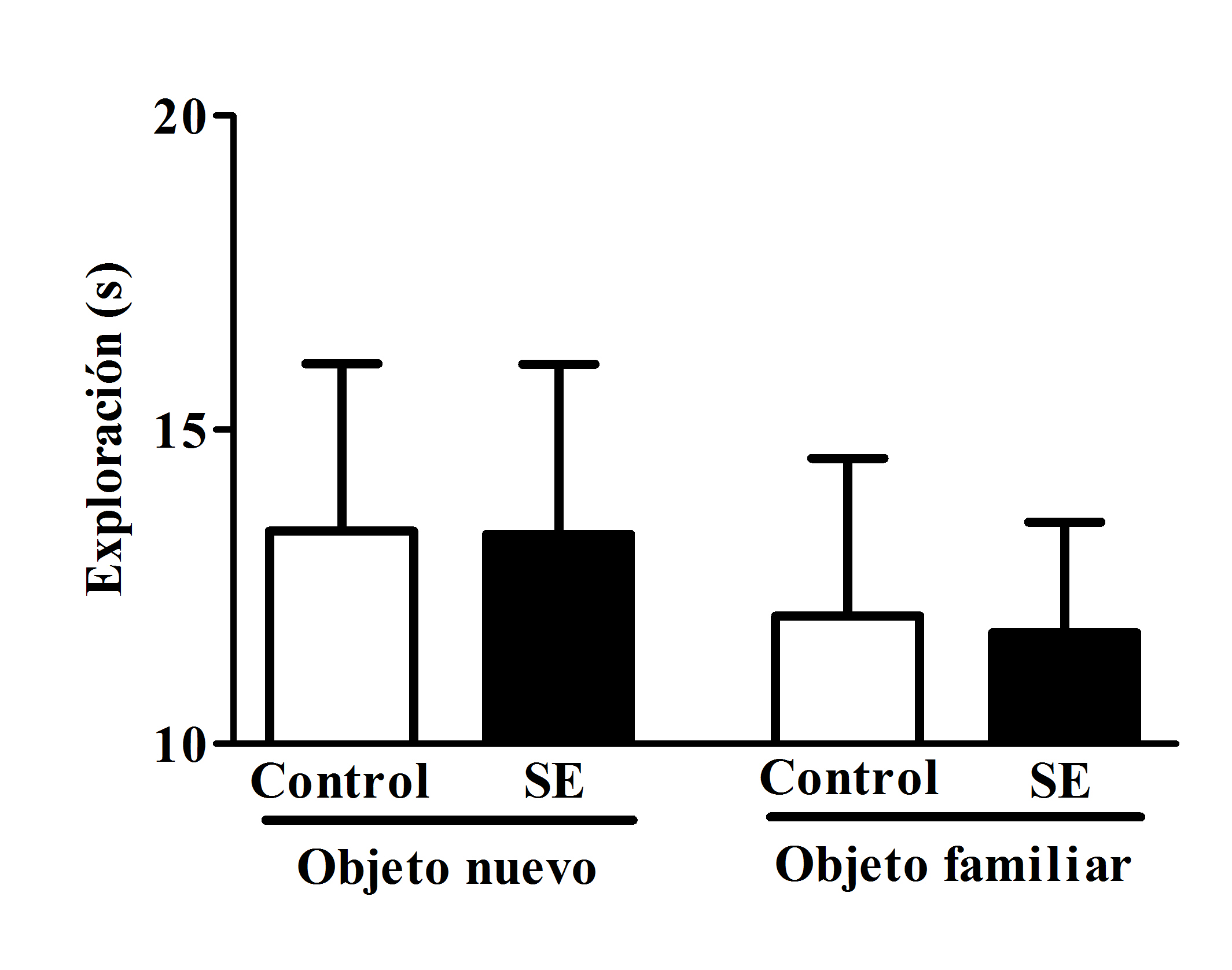
Figura 2. Tiempo promedio (media ± E.E.M.) de exploración del objeto nuevo y del objeto familiar en los grupos control y SE. No se detectaron diferencias estadísticamente significativas entre los grupos.
El valor de ID promedio del grupo de ratas con SE (3.6 ± 11.0 s) no resultó estadísticamente diferente al valor obtenido por los animales del grupo control (13.8 ± 14.2 s), aunque se observó una ligera disminución de este parámetro en los sujetos con SE (t=0.578, gl=16, p=0.571, figura 3).
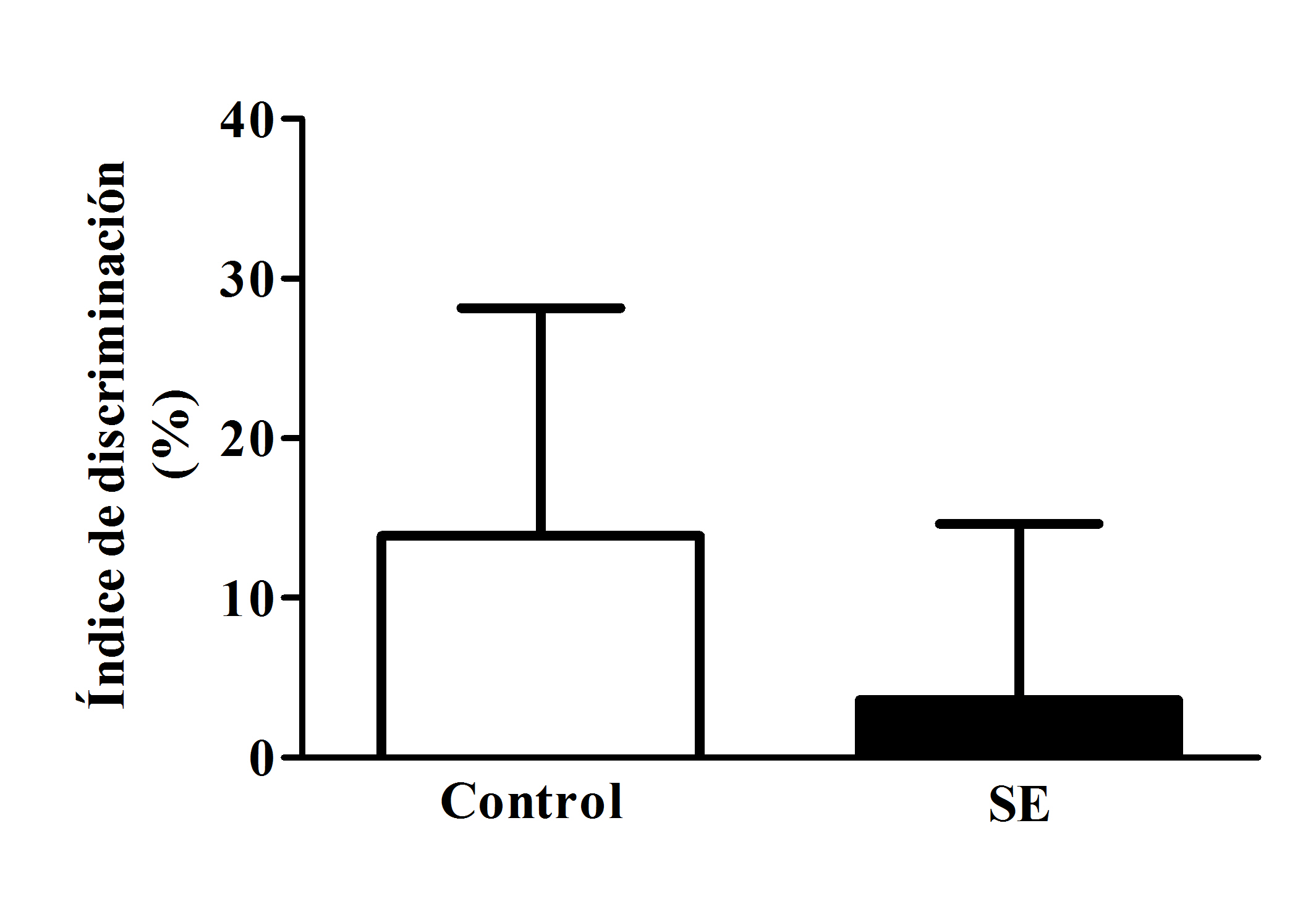
Figura 3. Índice promedio (media ± E.E.M.) de discriminación (ID) de los grupos control y SE obtenido en la prueba de reconocimiento de un objeto nuevo. No se detectaron diferencias estadísticamente significativas entre los grupos.
El analisis del ID individual de los animales del grupo SE y del grupo control, mostró que 60% (6/10) de las ratas que tuvieron crisis mostraron un valor negativo de ID, i.e., exploraron más el objeto familiar, mientras que sólo el 25% (2/8) de los animales control presentaron este comportamiento, aunque no resultó ser estadísticamente significativo (p=0.692) (Tabla 1).
GRUPO CONTROL |
|
GRUPO |
||
RATA |
ID (%) |
RATA |
ID (%) |
|
A4 |
76.923 |
A1 |
8.743 |
|
A5 |
-54.286 |
A2 |
64.303 |
|
A13 |
8.108 |
A3 |
-1.435 |
|
A14 |
40.123 |
A6 |
-36.641 |
|
A15 |
-0.319 |
A7 |
-7.353 |
|
A16 |
-19.298 |
A8 |
-10.836 |
|
A17 |
41.026 |
A9 |
-2.251 |
|
A18 |
18.719 |
A10 |
-49.123 |
|
Animales con ID negativo = 3/8 |
A11 |
20.27 |
||
A12 |
50.515 |
|||
Animales con ID negativo = 6/10 |
||||
No se detectaron diferencias en el número de animales con ID negativo entre los grupos experimentales (prueba exacta de Fisher, p=0.692).
Los hallazgos del estudio muestran que los animales que tuvieron SE convulsivo durante la segunda semana de vida, no muestran diferencias en la memoria de trabajo evaluada durante el día posnatal 21, en comparación con los sujetos control.
El modelo de reconocimiento de un objeto nuevo se implementó inicialmente para valorar la capacidad de la rata para reconocer un objeto familiar con el paso del tiempo, habilidad que se ha denominado memoria de reconocimiento.13 Este modelo aprovecha la naturaleza intrínseca de exploración de las ratas y se considera prometedor para detectar alteraciones mnemónicas pediátricas y de la adolescencia.14 Este modelo contempla dos funciones, la capacidad de detectar lo conocido (familiarity detection) y la capacidad de expresar esa función (recollection).15,16 Así, si los animales muestran una mayor interacción con el objeto novedoso durante la prueba (>50%), se puede presumir que han recordado el objeto muestra o familiar, lo que se considera como un indicativo de memoria.17 Por tal motivo este modelo explora posibles modificaciones en la memoria de trabajo (memoria a corto plazo).
Los resultados del presente proyecto, muestran que el tiempo de exploración del objeto nuevo y del objeto familiar de ratas que presentaron SE durante el día posnatal 14 es similar al de los animales que no convulsionaron o control. Este hecho también se refleja en el valor promedio del ID, ya que no fue diferente entre los grupos experimentales. Sin embargo, se registró la tendencia de valores de ID bajos en las ratas con SE respecto al grupo control, pero al realizar el contraste los resultados mostraron que aunque un mayor número de ratas del grupo SE presentaron un ID negativo, éste no resultó ser diferente al comprarse con el grupo control. La ausencia de diferencias estadísticamente significativas entre los tratamientos puede atribuirse a varios aspectos. Inicialmente, un hecho que podría enmascarar el efecto del SE en la memoria de trabajo podría estar relacionado con la baja capacidad de discriminación de objetos de las ratas próximas al destete (20-23 días de edad), según se propuso en un estudio previo.14 La explicación podría ser que si bien es cierto que los sujetos control exploran más el objeto nuevo que el objeto familiar (57% vs 43%, respectivamente), la diferencia es de menos de 2 s en promedio, i.e. el tiempo de exploración es cercano al 50:50. Se conoce que existen diferencias debido a la ontogenia en la capacidad de reconocer el objeto nuevo en este modelo experimental, al realizar la prueba 24 h después del periodo de entrenamiento,14 tal y como se realizó en este proyecto; en este reporte las ratas de tres semanas de edad presentaron un porcentaje de exploración del objeto nuevo por debajo del 50%.14 Por lo que es indispensable realizar un protocolo experimental similar al que se empleó en este trabajo, aunque analizando la memoria de trabajo a periodos más cortos después del SE, considerando que en este estudio el modelo de memoria se realizó 7 días después del SE. Debe recalcarse que se siguió este esquema experimental debido a que durante las primeras 48 h posteriores al SE los animales no se desenvuelven normalmente (algunas horas posteriores al SE los animales se encuentran hipoactivos, debido al desgaste que resulta de presentar convulsiones durante aproximadamente 5 h y al día siguiente se muestran hiperexcitables con una susceptibilidad aumentada a presentar una crisis convulsiva ante estímulos que originalmente no la desencadenarían) y a que requirieron de al menos cuatro sesiones diarias de habituación en la cámara de prueba (datos no mostrados de un estudio piloto). Asimismo, sería importante realizar la prueba máximo 1 h después de la pre-prueba o sesión de familiarización, con la finalidad de confirmar/descartar que el SE produce un efecto en la memoria indetectable a tiempos mayores, considerando lo observado por Reger y colaboradores.14 Como ya se mencionó anteriormente, el modelo de reconocimiento de un objeto nuevo resulta atractivo a la investigación porque no requiere una motivación externa o reforzamiento, además de que demanda periodos cortos de habituación y entrenamiento. Sin embargo, algunas de sus limitantes pueden ser los niveles bajos de exploración, el aumento de la actividad exploratoria al incrementar el tamaño de la arena, la preferencia hacia ciertos objetos que determinen la exploración que el roedor despliega hacia estos, cambios en la respuesta debido a diferentes intervalos o duración de las fases del modelo (i.e. habituación, familiarización (pre-pueba) y prueba), así como efecto del medio ambiente en la preferencia hacia uno de los objetos.18 Las consideraciones anteriores explicarían, en parte, la falta de evidencia experimental respecto a este tema en ratas en desarrollo. Por lo tanto, una perspectiva derivada de este trabajo, sería emplear modelos experimentales de memoria alternos, tal es el caso del laberinto acuático de Morris (LAM). Este modelo se ha empleado ampliamente para el estudio de las consecuencias de la epilepsia en procesos mnemónicos durante la etapa adulta.7-9 El LAM proporcionaría información sobre deterioro de la memoria y el aprendizaje sobretodo espacial, aunque la memoria de trabajo también podría ser explorada.
Desde el punto de vista neuroanatómico, el modelo de reconocimiento de un objeto nuevo involucra al lóbulo temporal importantemente,19,20 entre ellos al hipocampo21-24 y a la corteza perirrinal.25-27 Se sabe que el SE producido en ratas de 14 días de edad causa muerte neuronal extensa en el hipocampo, el tálamo y la amígdala,11, 28, 29 todas estas regiones ubicadas en el lóbulo temporal. Dado este efecto, hipotetizamos que el SE podría modificar la memoria de trabajo evaluada en el modelo de reconocimiento de un objeto nuevo. Sin embargo, los resultados señalan que el SE no causó deterioro en este proceso mnemónico a pesar de presentar una tendencia a hacerlo. Esto puede asociarse con la plasticidad cerebral/neuronal amplia que posee el cerebro en desarrollo,30 lo cual permite a las neuronas restantes suplir funcionalmente a las que se perdieron o que incluso otras regiones cerebrales del lóbulo temporal pudieran ejecutar las funciones que dejan de realizar las áreas que sufrieron muerte neuronal, en contraste con individuos adultos.
En resumen, un mayor número de ratas del grupo SE presentaron un ID negativo con respecto al grupo control, aunque la diferencia no fue estadísticamente significativa. Sin embargo, este resultado implicaría que el SE produce un deterioro en el reconocimiento de los objetos ya que la rata parece no recordar al objeto familiar. Considerando lo anterior, no podemos descartar que la ausencia de cambios puede deberse a que la muestra analizada no es suficiente para identificar el fenómeno.
El SE inducido a ratas de 14 días de edad no modifica la memoria de trabajo evaluada durante el día posnatal 21.
Al Consejo Nacional de Ciencia y Tecnología por el donativo de Investigación Básica otorgado a MLLM (número 106402) y la beca para la realización de estudios de doctorado otorgada a JSMM (número 223546).
- Engel J Jr. Report of the ILAE classification core group. Epilepsy 2006 47: 1558-68.
- Wasterlain CG y Chen J. Definition and classification of status epilepticus. En: Wasterlain CG y Treiman DM (Eds.) Status epilepticus: Mechanisms and management. The Mit Press 2006 pp 11-6.
- Hauser W. The prevalence and incidence of convulsive disorders in children. Epilepsia 1994 35:1-6.
- DeLorenzo RJ, Pellock JM, Towne AR, Boggs JG. Epidemiology of status epilepticus. J Clin Neurophysiol 1995 12: 316-25.
- DeLorenzo RJ, Hauser WA, Towne AR, Boggs JG, Pellock JM, Penberthy L, Garnett L, Fortner CA, Ko D. A prospective, population-based epidemiologic study of status epilepticus in Richmond, Virginia. Neurology 1996 46: 1029-35.
- Holmes GL, Ben-Ari Y. Seizures in the developing brain: perhaps not so benign after all. Neuron 1998 21: 1231-4.
- Stafstrom EC. Assessing the behavioral and cognitive effects of seizures on the developing brain. Prog Brain Res 2002 135: 377-90.
- Lynch M, Sayin U, Bownds J, Janumpalli S, Sutula T. Long-term consequences of early postnatal seizures on hippocampal learning and plasticity. Eur J Neuroscience 2000 12: 2252-64.
- Sayin U, Sutula TP, Stafstrom CE. Seizures in the developing brain cause adverse long-term effects on spatial learning and anxiety. Epilepsia 2004 45: 1539-48.
- Bear FM, Connors WB, Paradiso AM. Neuroscience: exploring the brain. Lippincott Williams and Wilkins, 2006 pp 726-35.
- Sankar R, Shin DH, Liu H, Mazarati A, Pereira de Vasconcelos A, Wasterlain CG. Patterns of status epilepticus-induced neuronal injury during development and long-term consequences. J Neurosci 1998 18: 8382-93.
- Wang YC, Huang CC, Hsu KS. The role of growth retardation in lasting effects of neonatal dexamethasone treatment on hippocampal synaptic function. PLos one 2010 5: e12806.
- Ennaceur A, Delacour J. A new one-trial test for neurobiological studies of memory in rats. 1: Behavioral data. Behav Brain Res 1988 31: 47-59.
- Reger ML, Hovda DA, Giza CC. Ontogeny of rat recognition memory measured by the novel object recognition task. Dev Psychobiol 2009 51: 672-8.
- Aggleton JP, Brown MW. Interleaving brain systems for episodic and recognition memory. Trends Cogn Sci 2006 10: 455-63.
- Fortin NJ, Wright SP, Eichenbaum H. Recollection-like memory retrieval in rats is dependent on the hippocampus. Nature 2004 431: 188-91.
- Akkerman S, Blokland A, Reneerkens O, van Goethem NP, Bollen E, Gijselaers HJ, Lieben CK, Steinbusch HW, Prickaerts J. Object recognition testing: Methodological considerations on exploration and discrimination measures. Behav Brain Research 2012 232: 335-47.
- Antunes M, Biala G. The novel object recognition memory: neurobiology, test procedure, and its modifications. Cogn Process 2012 13:93-110.
- Bachevalier J, Vargha-Khadem F. The primate hippocampus: ontogeny, early insult and memory. Curr Opin Neurobiol 2005 15: 168-74.
- Brown MW, Aggleton JP. Recognition memory: what are the roles of the perirhinal cortex and hippocampus? Nat Rev Neurosci 2001 2: 51-61.
- Baker KB, Kim JJ. Effects of stress and hippocampal NMDA receptor antagonism on recognition memory in rats. Learn Mem 2002 9: 58-65.
- Clark RE, Zola SM, Squire LR. Impaired recognition memory in rats after damage to the hippocampus. J Neurosci 2000 20: 8853-60.
- Hammond RS, Tull LE, Stackman RW. On the delay dependent involvement of the hippocampus in object recognition memory. Neurobiol Learn Mem 2004 82: 26-34.
- Vnek N, Rothblat LA. The hippocampus and long-term object recognition memory in the rat. J Neurosci 1996 8: 2780-7.
- Bussey TJ, Muir JL, Aggleton JP. Functionally dissociating aspects of event memory: the effects of combined perirhinal and postperirhinal cortex lesions on object and place memory in the rat. J Neurosci 1999 19: 495-502.
- Mumby DG, Pinel JP. Rhinal cortex lesions and object recognition in rats. Behav Neurosci 1994 108: 11-8.
- Winters BD, Forwood SE, Cowell RA, Saksida LM, Bussey TJ. Double dissociation between the effects of peri-postrhinal cortex and hippocampal lesions on tests of object recognition and spatial memory: heterogeneity of function within the temporal lobe. J Neurosci 2004 24: 5901-8.
- Kubova H, Druga R, Lukasiuk K, Suchomelova L, Haugvicova R, Jirmanova I, Pitkanen A. Status epilepticus causes necrotic damage in the mediodorsal nucleus of the thalamus in immature rats. J Neurosci 2001 21: 3593-9.
- Nairismägi J, Pitkänen A, Kettunen MI, Kauppinen RA, Kubova H. Status epilepticus in 12-day-old rats leads to temporal lobe neurodegeneration and volume reduction: a histologic and MRI study. Epilepsia 2006 47: 479-88.
- McCutcheon JE, Marinelli M. Age matters. Eur J Neurosci 2009 29: 997-1014.
| Recibido: 12 de enero de 2013 | Aceptado: 21 de febrero de 2013 |
Correspondencia:
Correspondencia: Centro de Investigaciones Cerebrales, Universidad Veracruzana Av. Dr. Luis Castelazo s/n, Col. Industrial Las Animas Xalapa, Ver. C.P. 91190. Teléfono: (228) 8418900. Ext. 13609 Fax: (228) 8418900. Ext. 13611 E-mail: leonorlopez@uv.mx
Este es un artículo de libre acceso distribuido bajo los términos de la licencia de Creative Commons, (http://creativecommons.org/licenses/by-nc/3.0), que permite el uso no comercial, distribución y reproducción en algún medio, siempre que la obra original sea debidamente citada.