La neurociencia del bienestar animal: teoría 80-20
- Inicio
- Comité Editorial
- Lineamientos
- Carta de Cesión de Derechos
- Información Legal
- Contacto
- ISSN 2007-3054
- Centro de Investigaciones Cerebrales
Universidad Veracruzana - Registrada en:
- Latindex
- Imbiomed
Articulo de Revisión
Genaro A. Coria-Avila1*, Deissy Herrera-Covarrubias2
Centro de Investigaciones Cerebrales, Universidad Veracruzana, Xalapa, Ver., México.1 Doctorado en Neuroetología, Universidad Veracruzana, Xalapa, Ver., México.2
Resumen/Abstract
Introducción
Conclusiones
Referencias
Correspondencia
Comúnmente se considera al bienestar animal cuando los animales están bien física y psicológicamente. Esto se logra cuando están libres: 1) de hambre, sed y malnutrición, 2) de incomodidad, 3) de dolor, 4) para expresar conducta normal, 5) de miedo y estrés. Este artículo tiene la intención de provocar al lector para reconsiderar el concepto de bienestar animal. La evidencia indica que el bienestar no es un estado constante, sino que debe ocurrir muchas veces al día. Se propone una teoría con la que argumentamos que el bienestar ocurre cuando la proporción de desear algo y obtenerlo es del 80-20%, respectivamente. Se discuten algunas bases neurobiológicas de las conductas motivadas que apoyan la nueva visión de bienestar.
Palabras clave: Dopamina, Opioides, Enriquecimiento ambiental, Bienestar, Deseo, Recompensa.
Animal welfare is commonly regarded as the physical and psychological well-being of animals, fulfilled if animals are free: 1) from hunger, thirst and malnutrition, 2) from discomfort, 3) from pain, 4) to express normal behavior, and 5) from fear and distress. This paper is meant to provoke the reader to re-think the concept of welfare. Evidence indicates that animal welfare is not a constant state, but rather it must be fulfilled several times a day. A theory is proposed arguing that well-being occurs when the proportion of desiring and obtaining something occurs in a 80-20% proportion, respectively. The neurobiological bases of motivated behaviors are discussed to support a new view on animal welfare.
Key words: Dopamine, Opioids, Environmental enrichment, Well-being, Desire, Reward.
Animal welfare is commonly regarded as the physical and psychological well-being of animals,1 and depending on the species it is inferred from behaviors and physiological responses that are believed to indicate normality.2-5 According to the Brambell's five freedoms,6 animals may experience well-being if they are free: 1) from hunger, thirst and malnutrition, 2) from discomfort, 3) from pain, 4) to express normal behavior, and 5) from fear and distress. The five tenets indicate that animals must have access to fresh water and to a diet so that they maintain full health and vigour. They must be provided an appropriate environment including shelter and a comfortable resting area. If needed, a rapid diagnosis and treatment must be available. Furthermore, facilities where they are kept must have sufficient space, and company of the animal's own kind to facilitate normal behavior. And finally, caretakers must ensure conditions and treatment to minimize mental suffering.
The main goal of this article is to provoke the reader to re-think the concept of welfare. Accordingly, we will discuss some ideas that suggest that animal welfare is not a constant state in which animals can stay throughout the day, but rather it must be fulfilled several times a day. We will discuss information on the neural bases of motivated behaviors to support the idea that well-being cannot be reached without experiencing a bit of hunger, discomfort, pain, fear or distress (minor aversive stimuli).7 In other words, we will argue that animal well-being is relative, and depends on the net positive state minus the negative state evoked by aversive stimuli. This is based on the fact that the brain in all species functions as a filter/integrator/modulator of environmental stimuli, so that animals can execute the best mental/behavioral output to cope with internal and external needs. The process of filtering/integrating/modulating environmental stimuli leads to desire and to execute behaviors indicative of such motivation. Desiring and obtaining something is part of a necessary sequence that we have arbitrarily set in a 80-20% proportion, respectively. Without desire a normally-rewarding stimulus may be experienced as neutral, or even aversive. This becomes really important in environmental enrichment programs which aim to increase desire, attention, and expectation in animals as part of their well-being.
2. Panksepp´s Circumplex of emotions
Panksepp, Knutson, and Burgdorf wrote a very interesting essay about how emotions function as signals that indicate potential increases or decreases in fitness.8 They argued that: “although animals are obviously not consciously computing their fitness, they are aware of their feelings at some level and respond accordingly, as evidenced by their behavior”. In the so-called affective circumplex they represent the whole repertoire of animal emotions into a circle, divided in four quadrants (Figure 1). The two quadrants on the right represent positive emotions (indicative of potential increases in fitness), whereas the two quadrants on the left represent negative emotions (indicative of potential decreases in fitness). In addition, the two higher quadrants are characterized by increased arousal, and the two lower quadrants represent decreased arousal. According to the authors, emotions that signal potential increases in fitness create a vector moving up and to the right, which generates positive feelings involving high arousal (e.g. euphoria, happiness, attention, high expectation). Likewise, removal of potential decrements in fitness creates a vector moving down and to the right, which generates positive feelings involving low arousal (e.g. relaxation, calmness, satiety). Potential decreases in fitness create a vector moving up and to the left, which generates negative feelings involving high arousal (e.g. anxiety, frustration), while removal of potential increments in fitness creates a corresponding vector moving down and to the left, which may generate different negative feelings involving low arousal (e.g. depression).
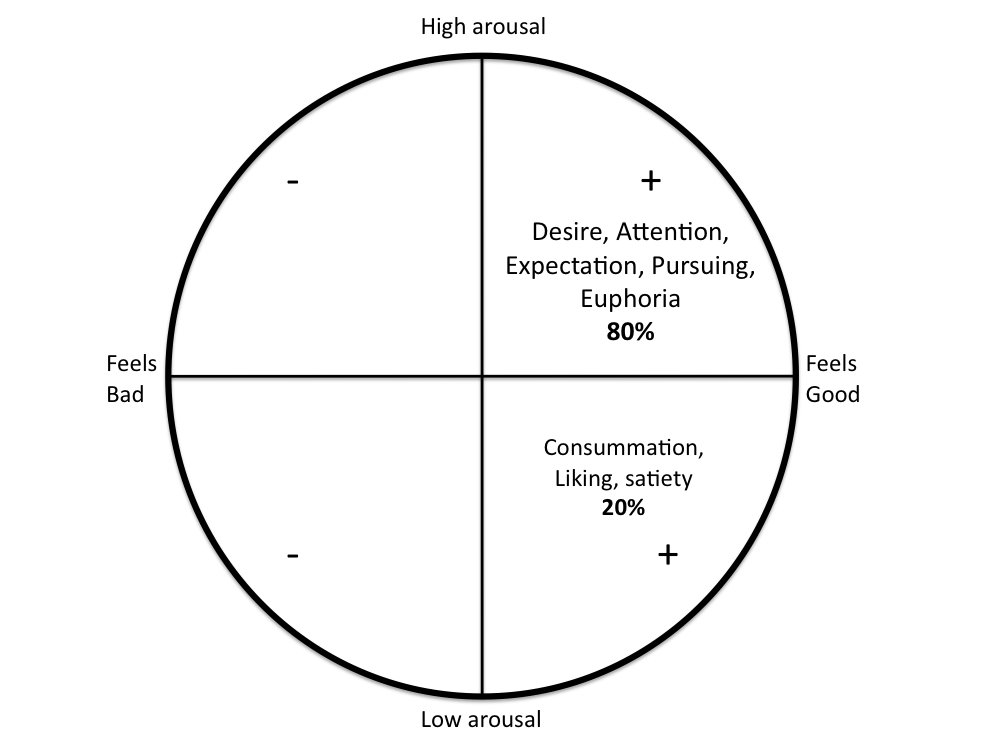
Figure 1. Modified from Panksepp´s circumplex of emotion adapted for our theory 80-20 of animal well-being. All animals experience positive or negative emotions produced by brain circuitries that evolved to signal increases or decreases in fitness. Accordingly, when animals are about to obtain something that increases fitness (e.g. food, sex, social companion) they may experience emotions that “feel good” with high arousal such as desire, and euphoria. A good amount of desire and euphoria (arbitrarily set here as 80% of the whole process of well-being) must occur before animals obtain the wanted stimulus. During consummation animals may experience emotions that “feel good” with low arousal, such as satiety, orgasm, relaxation, or calmness (arbitrarily set here as 20% of the whole process of well-being). Animal well-being is fullfilled when we provide the conditions to reach the upper right side of the circumplex first, and then the lower right. Ad libitum access to resources prevents animal well-being because it disrupts the normal neurological proccess of desire and consummation. Modified from circumplex of emotion of Panksepp, Knutson, and Burgdorf (2002).
The circumplex of emotions is extremely important for understanding animal welfare because well-being is experienced everytime an animal reaches the right quadrants. The logical sequence of motivated behaviors indicates that animals must be first in the right, upper quadrant so that they experience desire, attention and expectation of something that increases fitness as represented by potential reward, which makes them display behaviors to obtain the source of desire. They may expend sufficient amount of time in that quadrant (80% of time) before they obtain the reward and enjoy it (20% of time). This sequence of wanting something and obtaining the corresponding reward during consummation may be supported by neurocircuitries mediated by dopamine and opioids, respectively.
3. Proposal of theory 80-20
Yew-Kwang Ng9 described welfare as net happiness, or enjoyment minus suffering. Accordingly, all we would have to do to provide animal welfare is to increase “happiness” and to decrease “suffering”. In fact, the five freedoms are meant (in theory) to reach that goal. However, experimental data indicate that negative emotions found at the left side of the circumplex (evoked by stimuli that feel bad) may be necessary for the brain to compute a relative contrast when something positive is about to be obtained or has been obtained. In other words, being always on the right side of the circumplex is not possible by having free access to all resources or good stimuli, but rather by controlling access to them, which causes an increase of desire, expectation and attention, with the consequent enjoyment during consummation. Imagine for example a lost person in the forrest. He has been walking for several hours and he is running out of water and food. Finally, at the end of the day he finds his way to a village where people cheer him up and offer him a hot bowl of chicken soup. The state of euphoria caused by seeing the village lights after a long day of walking and sipping the soup when still hungry may be much higher compared to a full day of ad libitum access to soup and resting in a nice couch. Likewise, the state experienced during and after consummation may be experienced more intensively. Berry Spruijt defined welfare as the balance between positive (reward, satisfaction) and negative (stress) experiences or affective states,10 and very recently, Mellor indicated the relevance of the neural/cognitive foundations of motivational drives that energize and direct particular behaviours and their related subjective or emotional experiences.11
The theory 80-20 is somehow based on previous ideas of scientists dedicated to explore animal welfare.2-5,7,10,11 However, this theory specifically suggests that animal well-being is more easily experienced when an individual wants something before obtaining it. We have set it to 80% and 20%, respectively, keeping in mind that the proportion of wanting and desiring must outlast the proportion of consummation. Interestingly, indicators of “happiness” in humans show that it does not depend on how much we have, but rather on the relative degree in which a individual fulfills his/her expectations.12,13 The theory suggests that animal caretakers must provide conditions in which animals experience desire, and expectation of reward before they are allowed access for consummation. However, consummation (access to food, sex, companion, etc.) must eventually occur in order to avoid frustration. For example, studies on sexual behavior of laboratory rats have shown that experiencing the consummatory phase (i.e. ejaculation) without previous desire, expectation or increased attention does not induce reward. Pfaus argues that “rats that ejaculate with no preceding intromissions (as occurs after injections of the drug 8-OH-DPAT, a serotonergic drug that binds to presynaptic autoreceptors and blocks serotonin release) are said to have a facilitated ejaculation, but not a facilitated sexual motivation or desire”. Those rats fail to show copulatory conditioned place preference, which indicate that ejaculation that comes too quickly without former desire is not rewarding.14 By contrast, chronic treatment of male rats with the selective serotonin reuptake inhibitor fluoxetine (antidepressant) induces a progressive delay in the ejaculation latency in a timed test. Such treatment produces decreases in measures of sexual desire that are not secondary to decreases in motor ability. In other words, high levels of expectation of reward without consummation produce the eventual extintion of motivated behaviors and potential stereotypical behavior as well.
Thus, putting together the five freedoms and the theory 80-20 presented here, we would suggest that animal welfare (well-being) is reached when animals are free:
1) From hunger, thirst and malnutrition. However, easy ad libitum food and water is not the solution because animals do not have to work for it.
2) From discomfort. However, living in a shelter where nothing happens is not the solution because an environment with no conditions to desire, predict and expect may render animals bored and frustrated.
3) From pain. However, our knowledge about pain vs. nociception is limited in animals. Negative feelings or “feeling bad” may even occur in the abscence of physical punishment.
4) To express normal behavior. However, normal behavior in the wild is meant for coping with normals stressors and to obtain fitness. Normal behavior involves a wide repertoire of positive and negative emotions.
5) From fear and distress. However, fear and distress in a small proportion are needed to produce contrast, so that the brain identifies stimuli that may increase fitness, which is experienced as positive emotions.
4. Dopamine and desire
More than one decade ago Spruijt suggested a concept of welfare based on reward evaluating mechanisms in the brain.10 He argued that the balanced action of dopamine and opioids in the brain are responsible for the well-being of animals. The theory 80-20 proposed here suggests a proportion of action of dopamine and opioids, respectively. For example, the dopaminergic system is believed to mediate the appetitive fases of motivated behavior (wanting, desire, expectation, attention), and therefore it is important to understand the 80% proportion of animal welfare and well-being. The major systems for desire and expectation involve mesolimbic, nigrostriatal, and hypothalamic dopamine (DA) which control general attention to incentive stimuli (especially stimuli of learned significance), and responses made toward those stimuli. Neurons of the mesolimbic DA system arise in the midbrain (ventral tegmental area or VTA) and project diffusely to different limbic and cortical structures. For example they reach the amygdala, nucleus accumbens (NAcc or ventral striatum), olfactory tubercle and piriform cortex, lateral septum, and the anterior cingulate cortex (ACC). The mesocortical projections reach the medial prefrontal cortex (mPFC), important for executive control and inhibition of behavior. Neurons of the nigrostriatal system arise in the substantia nigra and project to the caudate and putamen, known collectively as the striatum.15
When DA is released it scapes the synaptic cleft to bind to extrasynaptic receptors and transporters.16,17 Such activity can be measured with different neurochemical techniques which depending on the sensitivity and time scale can measure tonic and phasic aspects of extracellular DA. Phasic DA is observed when it reaches high concentrations for periods of milliseconds. By contrast, tonic DA is observed over a period of several minutes or even hours. There is plenty of evidence indicating that both types of DA release are associated with desire, expectation, attention and prediction of rewards. For example, more that one decade ago it was shown that phasic DA release in primates responded to unexpected rewards (food), and to conditioned cues (a light) that predicted an inminent reward (food). In addition, that study showed that DA neurons decreased their phasic activity when the conditioned cues predicted reward but it was not delivered when expected.18 The study by Schultz indicated that DA activity was an important mechanism for the brain computation that mediates motivation, motivated behaviors and learning. Other former studies also showed that tonic DA is also elevated when animals detect unconditioned or conditioned cues that predict reward (i.e. sex).19-22
Physical or pharmacological manipulations of the different DA systems result in disruption of motivated behaviors. For example, many neuroleptic drugs used in veterinary medicine such as acepromazine, haloperidol, or azaperone function by blocking DA activity in the brain. Consequently, animals behave less motivated or even appear to be tamed. With low DA activity animals cannot focus on a given task, predict reward or pay attention to stimuli with normal incentive salience. By contrast, high levels of DA activity are associated with increased expectation, attention, and appetitive behaviors, which can lead to stereotypical behavior, pacing, aggression, self-mutilation and compulsive behaviors. Drugs like cocaine and amphetamines are known for increasing DA activity. However, those abnormal compulsive behaviors can also be observed in animals that are kept in poor environmental conditions with ad libitum access to food, and nothing else to predict or to get excited for. The theory 80-20 proposed here suggests that environmental enrichment for laboratory (when applicable), zoo or domestic animals must be focused on keeping an environment in which animals have the possibility to experience unexpected rewards (favourite snacks, companion), but also can experience desire, expectation and prediction of rewards with high levels of attention (e.g. finding food following olfactory cues; getting food out of a box; or working on a Skinner box-like device to get access to companion). Such circumstances may have a positive effect on the DA activity of animals to cover the 80% of animal well-being.
5. Opioids and reward
Desire is always linked to reward. In fact, many paradigms in psychology use appetitive behaviors as an indirect indicator of reward. For example, positive reinforcers are stimuli that increase the probability of ongoing behavior. Morsels of food to a hungry rat are positive reinforcers because they increase responding for them, and animals will show with behavior how rewarding the morsels are. However, large rewards, such as a big meal that induces satiety, produces a negative feedback on behavior by activating inhibitory neural pathways. Thus, it can be said that bad reinforcers do not increase behavior, good reinforcers increase behavior, and large reinforcers that produce satiety also inhibit behavior. In his concept of welfare Berry Spruijt also suggested opioids as a very important part of well-being.10 This is because opioids are believe to cause reward in the brain, and consequently are mediators of satiety of many motivated behaviors. However, other neurotransmitters such as serotonin, and endocannabinoids are also involved in reward and satiety. Many motivated behaviors are characterized by increased activity in the opioidergic, serotonergic, and endocannabinoid systems, and in some cases specific antagonist prevent satiety.14,23,24
The theory 80-20 argues that by obtaining and enjoying reward animals fulfill 20% of the whole experience of well-being. Opioids (and the other neurotransmitters involved in satiety) may be required after the long process of desire and wanting. Of course this is an oversimplification of the very complex process of motivated behaviors. In this simple explanation one can say that opioids bring animals to the right and lower quadrant of the circumplex of emotions, which produces calmness, relaxation, less anxiety, less fear, analgesia, and satiety (Figure 1). By contrast, the lack of opioids may be associated with hyperalgesia, anxiety, dysphoria, irritability, aggressive behavior, and stereotypic behaviors. Theories of addiction suggest that animals (and humans) find drugs extremely rewarding because they “hijack” neurocircuitries that would normally be activated when animals have found, pursued, and obtained something important for their fitness.25
6. Well-being as a cyclic 80-20 process
With this theory it is argued that by desiring (wanting something) and obtaining something animals experience the whole process of brain well-being. However, in a given day it would be impossible to keep an animal wanting something for 19 hrs and experiencing reward for 5 more. Rather, we suggest that the process must be repeated constantly during the day. Small rewards must occur after small efforts, and large rewards only after big efforts.
It is difficult, if not impossible, to indicate a universal rule of well-being applicable to all species. Consequently, we believe that the concept of 80-20 may be useful for domestic, zoo or wild animals. However, the reader must understand that this is a theory based on data from different species under different circunstances. The theory 80-20 reported here has arbitrarily set this proportion for the phase of desiring and obtaining something. It implies that a good program of environmental enrichment will keep animals bussy wanting something, and working out to obtaining it before they can actually enjoy it. Somehow it is very well understood that ad libitum access to resources is not advantageous to animal welfare, and rather produces disadvantages in the well-being of animals since they become bored, and may show unwanted behaviors indicative of a disbalance. Animal welfare programs must avoid easy access to food and rewards, or facilities that lack sufficient complexity for animals to “use their brain”. Thus, the programs must exercise intelligent access to food and expected (and also unexpected) stimuli that help animals to predict situations, pay attention, and get excited for a potential reward. Success for welfare programs may well depend on the balance between desire and reward.
- Duncan IJ. The welfare of farm animals: an ethological approach. Sci Prog 1987 71: 317-26.
- Korte SM, Olivier B, Koolhaas JM. A new animal welfare concept based on allostasis. Physiol Behav 2007 92: 422-8.
- Boissy A, Manteuffel G, Jensen MB, Moe RO, Spruijt B, Keeling LJ, Winckler C, Forkman B, Dimitrov I, Langbein J, Bakken M, Veissier I, Aubert A. Assessment of positive emotions in animals to improve their welfare. Physiol Behav 2007 92: 375-97.
- Yeates JW, Main DC. Assessment of positive welfare: a review. Vet J 2008 175: 293-300.
- Mendl M, Burman OH, Paul ES. An integrative and functional framework for the study of animal emotion and mood. P Roy Soc B-Biol Sci 2010 277: 2895-904.
- Farm Animal Welfare Council. Five freedoms, Press statement, UK Goverment (1979).
- Veissier I, Boissy A. Stress and welfare: two complementary concepts that are intrinsically related to the animal's point of view. Physiol Behav 2007 92: 429-33.
- Panksepp J, Knutson B, Burgdorf J. The role of brain emotional systems in addictions: a neuro-evolutionary perspective and new 'self-report' animal model. Addiction 2002 97: 459-69.
- Ng Y-K. Towards welfare biology: Evolutionary economics of animal consciousness and suffering. Biol Philos 1995 10: 255-285.
- Spruijt BM, van den Bos R, Pijlman FT. A concept of welfare based on reward evaluating mechanisms in the brain: anticipatory behaviour as an indicator for the state of reward systems. Appl Anim Behav Sci 2001 72: 145-171.
- Mellor DJ. Animal emotions, behaviour and the promotion of positive welfare states. N Z Vet J, 2012 60: 1-8.
- Matheny GL. Money not key to happiness, survey finds. Physician Exec 2008 34: 14-5.
- Kahneman D, Krueger AB, Schkade D, Schwarz N, Stone AA. Would you be happier if you were richer? A focusing illusion. Science 2006 312: 1908-10.
- Pfaus JG. Pathways of sexual desire. J Sex Med 2009 6: 1506-33.
- Gerfen CR. Functional neuroanatomy of dopamine in the striatum. En: Iversen L, Iversen SD, Dunnett SB, Björklund A Dopamine handbook. Oxford University Press, New York. 2010 pp 11-21.
- Garris PA, Ciolkowski EL, Pastore P, Wightman RM. Efflux of dopamine from the synaptic cleft in the nucleus accumbens of the rat brain. J Neurosci 1994 14: 6084-93.
- Smeets WJ, Gonzalez A. Catecholamine systems in the brain of vertebrates: new perspectives through a comparative approach. Brain Res Rev 2000 33: 308-79.
- Schultz W. Predictive reward signal of dopamine neurons. J Neurophysiol 1998 80: 1-27.
- Pfaus JG, Damsma G, Nomikos GG, Wenkstern DG, Blaha CD, Phillips AG, Fibiger HC. Sexual behavior enhances central dopamine transmission in the male rat. Brain Res 1990 530: 345-8.
- Pfaus JG, Damsma G, Wenkstern D, Fibiger HC. Sexual activity increases dopamine transmission in the nucleus accumbens and striatum of female rats. Brain Res 1995 693: 21-30.
- Pfaus JG, Phillips AG. Role of dopamine in anticipatory and consummatory aspects of sexual behavior in the male rat. Behav Neurosci 1991 105: 727-43.
- Wenkstern D, Pfaus JG, Fibiger HC. Dopamine transmission increases in the nucleus accumbens of male rats during their first exposure to sexually receptive female rats. Brain Res 1993 618: 41-6.
- Lorrain DS, Riolo JV, Matuszewich L, Hull EM.Lateral hypothalamic serotonin inhibits nucleus accumbens dopamine: implications for sexual satiety. J Neurosci 1999 19: 7648-52.
- Rodriguez-Manzo G, Fernandez-Guasti A. Opioid antagonists and the sexual satiation phenomenon. Psychopharmacology (Berl) 1995 122: 131-6.
- Bechara A. Decision making, impulse control and loss of willpower to resist drugs: a neurocognitive perspective. Nat Neurosci 2005 8: 1458-63.
| Recibido: 28 de agosto de 2012 | Aceptado: 16 de octubre de 2012 |
DVM, PhD. Genaro A. Coria-Avila. Centro de Investigaciones Cerebrales, Universidad Veracruzana Av. Dr. Luis Castelazo s/n, Col. Industrial Las Animas Xalapa, Ver. C.P. 91190 Teléfono: (52228) 8418900 Ext. 13609 Fax: (52228) 8418900 Ext. 13611 E-mail: gcoria@uv.mx
Este es un artículo de libre acceso distribuido bajo los términos de la licencia de Creative Commons, (http://creativecommons.org/licenses/by-nc/3.0), que permite el uso no comercial, distribución y reproducción en algún medio, siempre que la obra original sea debidamente citada.