Temporalidad en el incremento de receptores a prolactina en el hipotálamo de la rata macho durante la conducta sexual
- Inicio
- Comité Editorial
- Lineamientos
- Carta de Cesión de Derechos
- Información Legal
- Acerca de la Revista
- Bases de Datos
- Contacto
- ISSN 2007-3054
- Centro de Investigaciones Cerebrales
Universidad Veracruzana
Artículo de Investigación
Temporal increase of prolactin receptors at the hypothalamus of the male rat during sexual behavior
María Elena Hernández1,2, Martha Miquel3, Luz Irene Pascual1, Deissy Herrera1, Gonzalo Aranda-Abreu1,2, Leonor López1, Fausto Rojas1, Rebeca Toledo1,2, Jorge Manzo1
1Programa de Neurobiología, Universidad Veracruzana, Xalapa, Ver., México. 2Cuerpo Académico de Neuroquímica. 3Universidad Jaume I. Castellón. España.
Resumen/Abstract
Introducción
Material y Métodos
Resultados
Discusión
Conclusiones
Agradecimientos
Bibliografía
Correspondencia
Introducción: Tres grupos de neuronas dopaminérgicas del hipotálamo regulan a la adenohipófisis para liberar prolactina sistémica. La inhibición de dopamina estimula la liberación de prolactina, que a su vez se autorregula al estimular la liberación de dopamina. Esto es posible dado que las neuronas hipotalámicas tienen receptores a prolactina que además elevan su concentración cuando hay prolactina presente. En estudios previos mostramos que la conducta sexual masculina eleva los niveles sistémicos de prolactina, sugiriendo la hipótesis de que también modifica los receptores hipotalámicos.
Objetivo: analizar la fluctuación de los PRLr y su temporalidad en el hipotálamo de la rata macho durante eyaculaciones consecutivas.
Material y Métodos: Con ello, se organizaron dos experimentos utilizando ratas macho sexualmente expertas. En el primero, los machos copularon de una a tres eyaculaciones consecutivas, después se obtuvo el hipotálamo. En el segundo, se obtuvieron los hipotálamos a diferentes tiempos después de la eyaculación correspondiente. En ambos experimentos se cuantificaron los niveles del mRNA del receptor a prolactina mediante RT-PCR.
Resultados: Los primeros resultados mostraron que el nivel del receptor incrementa a partir de la segunda eyaculación y se mantiene al mismo nivel después de la tercera. El segundo experimento mostró incrementos del receptor 15 minutos después de una eyaculación con una elevación máxima entre los 30 y 40 minutos. Dos o tres eyaculaciones consecutivas dieron una curva similar, aunque con mayor intensidad.
Conclusiones: Esto sugiere que los niveles de prolactina durante la conducta sexual influyen sobre la temporalidad en el incremento de los receptores hipotalámicos. Así, se propone que el inicio de la conducta sexual dispara mecanismos para la regulación de prolactina y sus receptores hipotalámicos, con una fluctuación muy conservada en función del tiempo.
Palabras clave: Neuroendocrinología, RT-PCR, Eyaculación, Hormonas Peptídicas.
Introduction: Three groups of hypothalamic dopaminergic neurons regulate the adenohypophysis to release systemic prolactin. The inhibition of dopamine stimulates prolactin release that in turn is self-regulated by stimulating the release of dopamine. This is possible because dopaminergic neurons have prolactin receptors which increase their concentration in the presence of prolactin. In a previous study we showed that male sexual behavior increases the systemic level of prolactin, suggesting the hypothesis that also modifies their hypothalamic receptors.
Objetive: to analyze the fluctuation of prolactin receptors and its temporality in the hypothalamus of male rats following consecutive ejaculations.
Materials and Methods: Hence, two experiments were developed by using sexually experienced male rats. In the first, males copulated from one to three consecutive ejaculations, and the second, the hypothalamus was obtained at different times after the correspondent ejaculation. Then the hypothalamus was obtained to measure the level of mRNA of the prolactin receptor with RT-PCR.
Results: First results showed that the level of the receptor increases after the second ejaculation and remains elevated after the third. The second experiment showed increase of the receptors 15 minutes after one ejaculation with a maximal expression between 30 and 40 minutes. Two or three consecutive ejaculations gave a similar curve, but with a higher intensity.
Conclusions: This suggests that the level of prolactin during sexual behavior impacts the temporality in the increase of hypothalamic receptors. Thus, we propose that the start of sexual behavior triggers mechanisms for the regulation of prolactin and its hypothalamic receptors, with a conserved fluctuation as a function of time.
Keywords: Neuroendocrinology, RT-PCR, Ejaculation, Peptide Hormone.
El hipotálamo es una estructura de la que se tiene una gran cantidad de conocimiento, por lo que se sabe que cumple con múltiples funciones para la regulación de la homeóstasis. Dentro de ellas, se encuentra su papel como un área central que participa en el control neural de funciones reproductivas, fundamentalmente de tipo neuroendocrino. Entre estas se incluye su influencia sobre los lactotropos de la adenohipófisis para regular la liberación de prolactina (PRL) al torrente sanguíneo; control que realiza a través de tres tipos distintos de neuronas dopaminérgicas: las tuberoinfundibulares, las tuberohipofisiales y las periventriculares hipofisiarias,1 las primeras se localizan en el núcleo arcuato y las dos últimas en el núcleo periventricular.2
La secreción constante de dopamina (DA) por parte de estas neuronas, inhibe la liberación de PRL en los lactotropos. De tal manera que para que PRL sea liberada, estas neuronas hipotalámicas necesitan reducir la liberación de DA.2-4 Cuando se reducen los niveles portahipofisiarios de esta catecolamina, los lactotropos incrementan la liberación de PRL al torrente sanguíneo y a su vez, esta misma hormona activa un circuito de retroalimentación negativa al estimular nuevamente a las neuronas dopaminérgicas del hipotálamo para que vuelvan a liberar DA, con lo que se detiene la liberación de PRL.2, 5-7 La PRL estimula este circuito al unirse a sus receptores a PRL (PRLr) localizados en las neuronas dopaminérgicas.8-10 Con ello, se sabe que en este caso en particular el hipotálamo es una región que regula funciones reproductivas a través de un circuito corto entre la DA, la PRL y los PRLr (Fig. 1).
La concentración de PRLr y en particular, de su mRNA, es potenciada por su propio ligando, la PRL, mediante un mecanismo autocrino/paracrino de regulación a la alta.11 Pero existen también otros mecanismos asociados a diferentes situaciones fisiológicas con otros ligandos que también contribuyen a la elevación del receptor, como situaciones de embarazo y lactancia.12-13 De esta información se desprende el hecho de que la gran mayoría de estudios sobre estos procesos de regulación se han realizado en hembras. Sin embargo, esta relación fisiológica en machos es menos entendida. En un estudio previo realizado en ratas macho mostramos que la PRL tiene un nivel basal en sangre, con dos picos de incremento en las transiciones del día a la noche y de la noche al día, y que la conducta sexual en una serie copulatoria con varias eyaculaciones consecutivas, dispara un incremento en los niveles sistémicos de PRL, con la máxima expresión después de la segunda eyaculación, a partir de la cual los niveles empiezan a descender.14
Estos hallazgos sobre las fluctuaciones en la concentración sanguínea de PRL durante la conducta sexual masculina, llevaron a proponer la hipótesis de que los cambios sistémicos de PRL estarían estimulando también fluctuaciones en la concentración de PRLr en las neuronas hipotalámicas. Por ello, el objetivo en el presente estudio fue estudiar la fluctuación de los PRLr y su temporalidad en el hipotálamo de la rata macho durante eyaculaciones consecutivas.
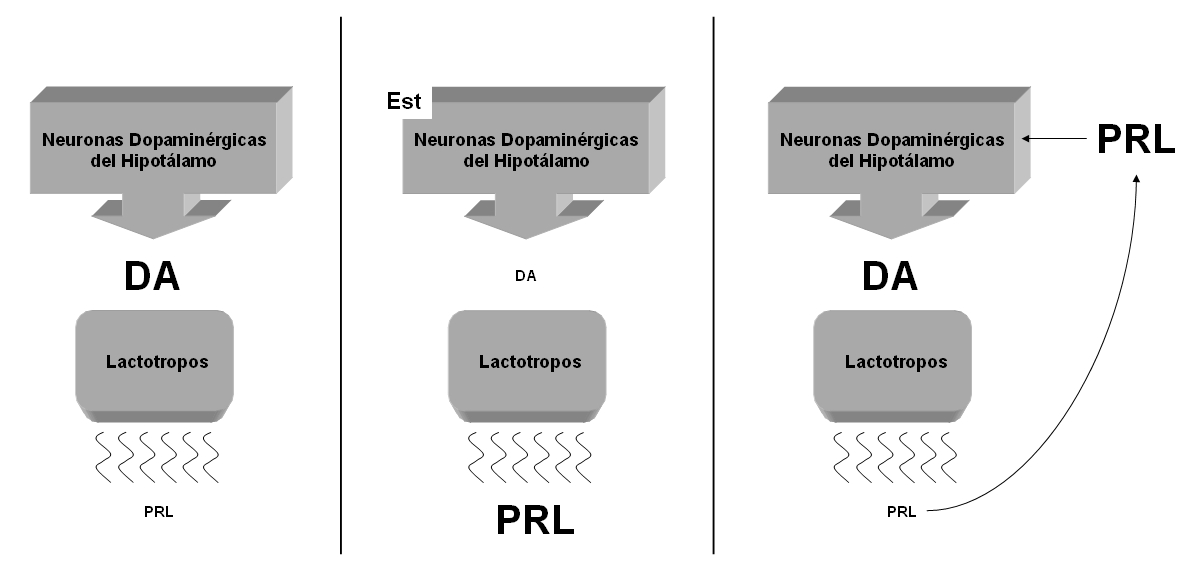
Figura. 1. La liberación de dopamina (DA) por las neuronas del hipotálamo inhibe la secreción de prolactina (PRL) de los lactotropos de la adenohipófisis (imagen izquierda). La presencia de un estímulo (Est) que reduce la actividad de las neuronas dopaminérgicas inhibiendo la liberación de DA, estimula a los lactotropos para que liberen PRL (imagen central). La PRL liberada estimula a su vez a las neuronas dopaminérgicas para que vuelven a liberar DA y para que reduzcan la liberación de PRL (imagen derecha). El tamaño de las letras DA y PRL indican una mayor o menor liberación.
2.1 Sujetos
Se utilizaron ratas macho adultas, sexualmente expertas, de la cepa Wistar (200-300 g de peso corporal). Para las pruebas de conducta sexual se utilizaron hembras ovariectomizadas y tratadas con hormonas esteroides para inducirles receptividad en el momento de las pruebas. Así, las hembras se inyectaron subcutáneamente con benzoato de estradiol (10 µg) y progesterona (500 µg) 48 y 4 horas antes de las pruebas respectivamente. Todas las ratas se mantuvieron en cajas de acrílico (50 x 30 x 20 cm; cinco animales del mismo sexo por jaula), que tenían una cama de aserrín constantemente limpio y se les proveía de agua y alimento (Harlan México) ad libitum. El alojamiento de las jaulas fue en un cuarto con ciclo luz-oscuridad invertido de 12-12 hrs (el apagado de la luz era a las 8:00 hrs). El proceso experimental se realizó respetando los estándares marcados por Norma Oficial Mexicana (NOM-062-ZOO-1999) y por las Políticas para el Uso de Animales en Neurociencias de la Society for Neuroscience.
2.2 Experimento 1
Este experimento tuvo el propósito de determinar la fluctuación del mRNA del PRLR en el hipotálamo de machos que ejecutaron de 1 a 3 eyaculaciones consecutivas. Para ello se utilizaron un total de 18 machos. Cada macho se colocó en una arena de conducta sexual en donde se introdujo a una hembra receptiva para iniciar la cópula. A partir de ahí se organizaron tres grupos de animales, el grupo 1 fue de sujetos que después de eyacular una vez, se tomaron de la arena de conducta, se sacrificaron y el hipotálamo fue extraído para su posterior análisis (n= 6); en el grupo 2, los sujetos se dejaron en la arena con la hembra hasta alcanzar dos eyaculaciones consecutivas, posteriormente se hizo el procedimiento para la obtención del hipotálamo (n= 6); y a los sujetos del grupo 3 se les dejó en la arena hasta que alcanzaron tres eyaculaciones consecutivas, para después extraerles el hipotálamo. Los resultados mostraron que una tercera eyaculación ya no produce un mayor incremento en el mRNA del PRLr, por lo que surgió la pregunta de cuál es la temporalidad en la expresión de este mensajero después de las eyaculaciones, lo que se respondió en el siguiente experimento.
2.3 Experimento 2
Aquí se utilizaron un total de 108 machos, organizados en tres grupos (n=36 cada uno). A los del grupo 1 se les permitió tener una eyaculación, posteriormente los hipotálamos de los sujetos (n= 6 en cada caso) fueron obtenidos a seis tiempos variables: inmediatamente después de la eyaculación, 5 minutos después, 15 minutos después, 30 minutos después, 60 minutos después y 120 minutos después. A los del grupo 2 se les dejó que tuvieran dos eyaculaciones y los hipotálamos se obtuvieron después de pasar los mismos tiempos anteriores. A los del grupo 3 se les dejó alcanzar tres eyaculaciones y se repitió el procedimiento.
2.4 Procesamiento de Hipotálamos y RT-PCR
Los hipotálamos fueron homogeneizados en 600 µl (1ml/100mg) de Solución D (Tiocianato de guanidium 4M, Citrato de Sodio 7.25mM, n-Lauroyl-Sarcosina 0.5%, ß-Mercaptoetanol 300µl/50ml). Posteriormente se adicionaron 100 µl de Acetato de Sodio 3M (pH 5.2), 1 ml de Fenol (balanceado con Tris pH 8.8) y 200 µl de Solución Savage (Cloroformo con Alcohol Iso-amil en proporción 24:1). La mezcla se colocó en hielo por 15 min y se centrifugó por 20 min a 12000 rpm a 4°C. La fase acuosa se precipitó adicionando un volumen de Isopropanolol por 4 hrs a -70°C. La pastilla de RNA se obtuvo por centrifugación (30 min a 12000 rpm y 4°C), se lavó con Etanol al 70% y se disolvió en agua tratada con DEPC. El RNA se cuantificó por mediciones de densidad óptica a 260 nm.
El mRNA del PRLr se amplificó con la técnica de Reacción en Cadena de la Polimerasa por Transcripción Reversa (RT-PCR, por sus siglas en inglés), utilizando un primer sentido 5’-CTCCTGACAAGGAAACATTCACCTGCTG-3’ (nucleótidos, nt, 305-333, 28mer) y primer antisentido 5’-TCAACGATGTAAGTCACATCCACATAA-3’ (NT 543-570, 27MER).
2.5 Análisis Estadístico
Los datos se analizaron mediante un Análisis de Varianza (ANOVA), seguida de la prueba post hoc de Dunnett, cuando el valor F resultó significativo F<0.05.
3.1 Experimento 1
Los datos mostraron que los niveles de mRNA del PRLr en el hipotálamo incrementaron significativamente después de eyaculaciones consecutivas [F(3,31)= 87.25, p<0.01]. Después de la primera eyaculación no se observó ningún cambio, pero el incremento se detectó después de la segunda eyaculación, manteniéndose la misma elevación después de tres eyaculaciones (Fig. 2).
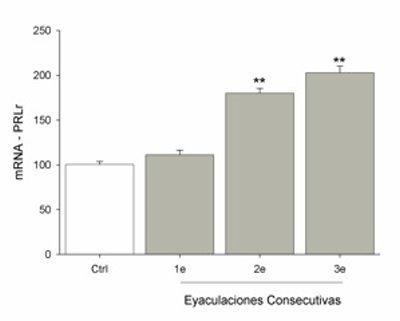
Figura 2. El incremento del RNA mensajero (mRNA) de los receptores a prolactina (PRLr) se observa a partir de una segunda eyaculación consecutiva, manteniéndose elevado después de una tercera eyaculación, aunque al mismo nivel.
3.2 Experimento 2
Los resultados del Experimento 1 dieron pie a plantear si el incremento similar del mRNA del PRLr después de dos o tres eyaculaciones consecutivas era producto de las eyaculaciones o de una función del tiempo en el que el macho estaba ejecutando la conducta. Para ello, se tomó el tiempo promedio que los machos utilizaron para ejecutar eyaculaciones consecutivas (Tabla 1) y se obtuvieron los resultados de este experimento. Así, se observaron incrementos significativos en el nivel del mensajero cuando los machos ejecutaron una eyaculación [F(5,35)= 30.98, p<0.01]; la medición a diferentes tiempos mostró un aumento significativo a partir de los 15 minutos, llegando alcanzar un nivel máximo a los 30 y 60 minutos, regresando después a niveles basales (Fig. 3). Los machos que ejecutaron dos eyaculaciones también dieron resultados significativos [F(5,35)= 174.80, p<0.01]; la medición a diferentes tiempos mostró el máximo aumento desde los 5 y hasta los 30 minutos, observándose un descenso a los 60 minutos y regresando a valores basales después de los 120 minutos (Fig. 3). La ejecución de tres eyaculaciones consecutivas también dieron resultados significativos [F(5,35)= 65.38, p<0.01], aunque en este caso la máxima expresión se observó 5 minutos después, empezando a descender a los 15 minutos y regresando a valores basales a partir de los 30 minutos (Fig. 3).
El traslape de estos datos en función del tiempo mostró que la ejecución de la conducta sexual con una o más eyaculaciones consecutivas tiene un patrón establecido en la elevación del mRNA del PRLr en el hipotálamo, donde se observa el inicio del incremento a partir del minuto 15 de que el macho está en contacto con la hembra, que finaliza después de los 60 minutos independientemente del número de eyaculaciones (Fig. 4). La máxima expresión de receptores se da alrededor de los 30 y 40 minutos después de iniciada la interacción sexual, y una segunda y tercer eyaculación consecutiva sólo elevan el nivel de receptores en ese momento, pero el pico de expresión y la curva de temporalidad de la expresión se mantuvieron constantes (Fig. 4).
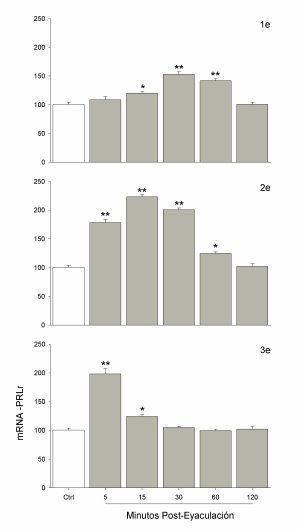
Fig. 3. El incremento del RNA mensajero (mRNA) de los receptores a prolactina (PRLr) muestra una temporalidad diferente después de cada una de las eyaculaciones consecutivas. Entre más eyaculaciones ejecuta el macho, más rápido se ven los incrementos y el descenso a niveles basales.
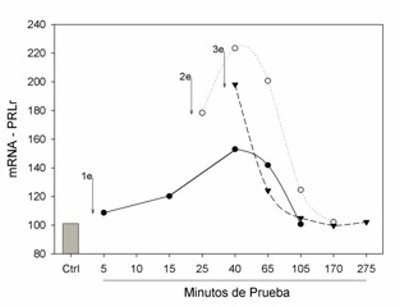
Figura 4. El incremento del RNA mensajero (mRNA) de los receptores a prolactina (PRLr) muestra una temporalidad que depende del tiempo en que inicia la primera interacción entre el macho y la hembra. Se observa un pico en la expresión del mensajero entre los 30 y 40 minutos después del primer contacto con la hembra, independientemente del número de eyaculaciones. Una segunda o tercer eyaculación incrementa el nivel del receptor pero en el mismo tiempo, lo que muestra una temporalidad bastante marcada en el incremento del receptor.
Previamente mostramos que la ejecución de la conducta sexual masculina con eyaculaciones repetidas produce un incremento transitorio en los niveles sistémicos de PRL en la rata macho14 y en este trabajo mostramos que una temporalidad similar ocurre en los PRLr en el hipotálamo. Esto sugiere que los niveles de PRL durante la conducta sexual tienen un impacto directo sobre la temporalidad en el incremento de los PRLr hipotalámicos. A pesar de ello, el primer eslabón en esta cadena de eventos estaría en la reducción de la liberación de DA por las neuronas hipotalámicas. En otros sistemas dopaminérgicos, tales como el nigroestriado, el mesolímbico y el área preóptica media, se observa el caso contrario en donde se estimula la liberación de DA durante la conducta sexual masculina en respuesta al incremento en los niveles de testosterona,15 que ya también mostramos previamente que tiene una elevación significativa durante eyaculaciones consecutivas.16 Sin embargo, es importante destacar el reporte de otros autores con respecto a las neuronas tuberoinfundibulares, donde la testosterona les reduce su actividad,17 sugiriendo la reducción en la liberación de DA y, por consiguiente, la liberación de PRL. Con ello, la testosterona parece ser la iniciadora del proceso de liberación de PRL al reducir la actividad de las neuronas dopaminérgicas y consecuentemente la síntesis de PRLr.
La PRL liberada reactiva a su vez a las neuronas dopaminérgicas para que liberen nuevamente DA.1 Sin embargo, esta estimulación no es constante, dado que la concentración de PRL en suero es temporal, independientemente del número de eyaculaciones ejecutadas.14 Aquí estamos reportando que la concentración del mRNA para el PRLr tiene un patrón similar. Independientemente del número de eyaculaciones, el receptor se eleva después de 30-40 minutos que el macho y la hembra han empezado a interaccionar. La diferencia entre eyaculaciones consecutivas es que si el macho ejecuta una segunda o tercer eyaculación, la concentración se eleva significativamente, pero en el mismo periodo en el tiempo, esto es, entre los 30 y 40 minutos. Esta información muestra que el inicio de la conducta sexual dispara mecanismos para la liberación y regulación tanto de PRL como de sus receptores hipotalámicos, y que estos mecanismos tienen una periodicidad muy conservada, que es intensificada pero no modificada por eyaculaciones consecutivas. Con esta información se propone que la fluctuación temporal tanto de PRL como del PRLr en el hipotálamo son procesos de corta duración indispensables para disparar otros mecanismos que regulan a la conducta sexual masculina. Esta observación no es propia de la PRL, dado que se sabe también que en hembras la exposición breve a picos de hormonas esteroides es suficiente para disparar los mecanismos que llevan a la conducta de lordosis.18 Así, el incremento temporal observado en los niveles del PRLr del hipotálamo pudiera servir para disparar distintos procesos fisiológicos en las neuronas dopaminérgicas, que ya iniciados no requieren de mayor activación por los receptores, lo que iniciaría su desaparición. Queda ahora resolver cual es el mecanismo fisiológico que regresa a los PRLr a sus niveles basales a pesar de que la conducta sexual sigue activa.
Esto sugiere que los niveles de prolactina durante la conducta sexual influyen sobre la temporalidad en el incremento de los receptores hipotalámicos. Así, se propone que el inicio de la conducta sexual dispara mecanismos para la regulación de prolactina y sus receptores hipotalámicos, con una fluctuación muy conservada en función del tiempo.
Proyecto Conacyt No. 106531 (Dra. María Elena Hernández), Cuerpo Académico de Neuroquímica UV-CA-137; Programa de Incorporación de Investigadores Españoles en Universidades Mexicanas AECI 2008 (Dra. Martha Miquel); Beca de la Secretaria General de Universidades del Ministerio de Educación, España SAB2009-0159 (Dr. Jorge Manzo Denes); Becas Conacyt No. 35258 (Deissy Herrera) y Beca Conacyt No. 209821 (Luz Irene Pascual).
- Demaria JE, Lerant AA, Freeman ME. Prolactin activates all three populations of hypothalamic neuroendocrine dopaminergic neurons in ovariectomized rats. Brain Res 1999 837:236-241.
- Ben Jonathan N, Hnasko R. Dopamine as a prolactin (PRL) inhibitor. Endocr Rev 2001 22:724-763.
- Hernandez ME, del Mar HM, Diaz-Munoz M, Clapp C, de la Escalera GM. Potentiation of prolactin secretion following lactotrope escape from dopamine action. II. Phosphorylation of the alpha (1) subunit of L-type, voltage-dependent calcium channels. Neuroendocrinology 1999 70:31-42.
- del Mar HM, Garcia Ferreiro RE, Garcia DE, Hernandez ME, Clapp C, Martinez dlE. Potentiation of prolactin secretion following lactotrope escape from dopamine action. I. Dopamine withdrawal augments l-type calcium current. Neuroendocrinology 1999 70:20-30.
- Ben Jonathan N. Dopamine: a prolactin-inhibiting hormone. Endocr Rev 1985 6:564-589.
- Freeman ME, Kanyicska B, Lerant A, Nagy G. Prolactin: structure, function, and regulation of secretion. Physiol Rev 2000 80:1523-1631.
- Lamberts SW, Macleod RM. Regulation of prolactin secretion at the level of the lactotroph. Physiol Rev 1990 70:279-318.
- Lerant A, Freeman ME. Ovarian steroids differentially regulate the expression of PRL-R in neuroendocrine dopaminergic neuron populations: a double label confocal microscopic study. Brain Res 1998 802:141-154.
- Pi XJ, Grattan DR. Increased prolactin receptor immunoreactivity in the hypothalamus of lactating rats. J Neuroendocrinol 1999 11:693-705.
- Pi XJ, Grattan DR. Increased expression of both short and long forms of prolactin receptor mRNA in hypothalamic nuclei of lactating rats. J Mol Endocrinol 1999 23:13-22.
- Swaminathan G, Varghese B, Fuchs SY. Regulation of prolactin receptor levels and activity in breast cancer. J Mammary Gland Biol Neoplasia 2008 13:81-91.
- Augustine RA, Kokay IC, Andrews ZB, Ladyman SR, Grattan DR. Quantitation of prolactin receptor mRNA in the maternal rat brain during pregnancy and lactation. J Mol Endocrinol 2003 31:221-232.
- Bole-Feysot C, Goffin V, Edery M, Binart N, Kelly PA. Prolactin (PRL) and its receptor: actions, signal transduction pathways and phenotypes observed in PRL receptor knockout mice. Endocr Rev 1998 19:225-268.
- Hernandez ME, Soto-Cid A, Rojas F, Pascual LI, Aranda-Abreu GE, Toledo R et al. Prostate response to prolactin in sexually active male rats. Reprod Biol Endocrinol 2006 4:28.
- Hull EM, Du J, Lorrain DS, Matuszewich L. Testosterone, preoptic dopamine, and copulation in male rats. Brain Res Bull 1997 44:327-333.
- Hernandez ME, Soto-Cid A, Aranda-Abreu GE, Diaz R, Rojas F, Garcia LI et al. A study of the prostate, androgens and sexual activity of male rats. Reprod Biol Endocrinol 2007 5:11.
- Toney TW, Lookingland KJ, Moore KE. Role of testosterone in the regulation of tuberoinfundibular dopaminergic neurons in the male rat. Neuroendocrinology 1991 54:23-29.
- Green R, Luttge WG, Whalen RE. Induction of receptivity in ovariectomized female rats by a single intravenous injection of estradiol-17. Physiol Behav 1970 5:137-141.
| Recibido: 7 de octubre de 2010 | Aceptado: 7 de diciembre de 2010 |
Correspondencia:
Correspondencia: Dra. María Hernández Aguilar, Programa de Neurobiología, Universidad Veracruzana, Xalapa, Ver., Tel.: (228) 841-8900 Ext. 13615, Correo: elenahernandez@uv.mx
Este es un artículo de libre acceso distribuido bajo los términos de la licencia de Creative Commons, (http://creativecommons.org/licenses/by-nc/3.0), que permite el uso no comercial, distribución y reproducción en algún medio, siempre que la obra original sea debidamente citada.